|
|
第9章 ハエからカビへ
ビードルとテータムがvの可能性が高い物質を一度は結晶化できたのだから、その化学構造はすぐに解決できると期待したのも無理はなかった。しかしエディンバラに着いた時に自分達が知らないうちに他の研究者に出し抜かれていたことを彼らは知った。スジコナマダラメイガ(Ephestia)の眼色についてアルフレッド・キューンと共同研究をしていた有名なドイツ人化学者のアドルフ・ブテナントが、細菌の働きによってトリプトファンからv物質が生産される観察を記述したテータムの論文1をおそらく読んで、v物質の作用を決定するために様々なトリプトファンの代謝中間体を試験していたのだった。彼らは哺乳類におけるトリプトファンの最初の分解産物として同定されていたキヌレニンがショウジョウバエとスジコナマダラメイガの眼色に関する突然変異体の眼色を正常に回復することを示した2。テータムはキヌレニンの同定に失敗したが、それは彼がバチルス菌の培養物から得た結晶がショ糖と結合したキヌレニンだったからである。テータムは、ショ糖を取り除いてもv物質の活性が失われないことを見て、すぐに得られた物質は実際にドイツ人グループが報告した本物のキヌレニンであることを確認した3。100分の1ミリグラムという少量のキヌレニンをバーミリオンの蛹に注入すると羽化したハエで正常な眼色の回復が認められた。
ビードルはアイオワ州立大学のクラレンス P.・バーグ教授に真性キヌレニンの提供を依頼した。バーグはキヌレニンの当初の同定に関与した人物で、彼のキヌレニンはテータムが分離した物質と化学的に同一だった。実は、1947年1月9日のワフー民主党新聞に書かれたビードルに関する記事を読んだバーグは、そこでビードルがワフーで育った有名な人物だったことを初めて知ったのだった。バーグの家族は1930年代にワフーに定住し、馬具職人だった父はビードル農場とビードルの父をおそらく知っていただろう4。キヌレニンが蓄積すると考えられるシンナバー突然変異体の蛹の組織から直接キヌレニンが分離されたことで、v物質の化学的性質が最終的に決定づけられた5。cn物質が当時はまだ知られていなかったキヌレニンの代謝物質3-ヒドロキシキヌレニンであることが分かったのは1949年のことだった6。
ビードルとエフルッシがショウジョウバエのホルモンだと信じた物質は昆虫に特有ではなく、むしろ細菌でも哺乳動物でもトリプトファンの正常な分解過程で作られる物質だったことが分かった。今日では、バーミリオン遺伝子はトリプトファンをN-フォルミルキヌレニンに変換する酵素をコードすること、さらに別の酵素がN-フォルミルキヌレニンをキヌレニンに変換することが分かっている。シンナバー遺伝子がコードするのは、キヌレニンを3-ヒドロキシキヌレニンに変換する酵素である。対応する突然変異体はこれらの酵素活性を欠いている7。
ブテナントの3-ヒドロキシキヌレニンに関するその後の研究で、ふたつの分子が結合した後でそれがさらに重合して茶色の色素ができること、それぞれの段階は遺伝的に特定された酵素によって触媒されることが分かった8。対照的に、赤色色素を形成する一連の反応については、その反応経路をブロックする数多くの突然変異が見つかっているにもかかわらず、まだほとんど何も分かっていない。モルガンの有名な白眼のハエをもたらすホワイト突然変異では、おそらく赤色と茶色の両色素の形成に共通なある段階がブロックされていることで、どちらの色素も形成できないのだろう。
v、cn物質の化学的同定の最初の成功者を目指して5年を費やした努力が報われなかったことにビードルとエフルッシは失望した。ドイツ人グループによるv物質がキヌレニンだという発見は、ビードルとテータムの発見が鍵だったとするスクープがあったことで、彼らの失望感は一層我慢がならないものになった。パリとスタンフォードの研究仲間も二人の努力が無に帰したと感じた。ビードルは、ブテナントの行動は科学者に相応しくない、もっと言えば 「意地が悪い」 と考え不愉快に思った。しかし、彼らが混乱したのは明らかだが、自分達の発見をv物質同定の踏み石にしたブテナントの行動は非倫理的だったとビードルが信じたかどうかは明らかではない。ブテナントはそれ以前に既にトリプトファン様の化学物質の研究に従事していたのであって、キューンのグループとの共同研究はテータムが与えた鍵となる事実を知ったからだけではなく、二人が共同すればその物質を即座に試験することができると考えたからだった。ビードルとエフルッシのグループがリプトファンの様々な化学的派生物質の試験に失敗したのは何故だろうと人は考えるかも知れない。彼らはトリプトファンそのものがv物質ではないこと、細菌によって活性のある物質に変換される何かであることを知っていた。おそらく彼らはトリプトファンの代謝で作られる化学物質がv物質である可能性を十分吟味することができなかったか、あるいはそうした化学物質を知らなかったか調べる術をもたなかったのであろう。
エフルッシがどのようにしてブテナントの発見を知ったのか、それを知ってどのように反応したかについてはよく分からない。ドイツ人達の論文が発表されたとき、エフルッシは科学よりは家族をフランスから避難させる方策を探すことに集中していた。エフルッシはエディンバラからパリに戻るとすぐに部署替えを命じられた。家族がオルレアンに留まる間は近くの田舎にあった実験室に勤務することになり、そこでショウジョウバエとは全く異なる仕事を任された。オルレアンにいる間に彼はビードルに頼んで、パリへ秘密でときどき出かける際に読むために論文のリプリントや科学ニュースをパリの実験室へ送ってもらった9。次の半年間は週に一度パリを訪ねることができたが、実験室はいつも人影がなくもぬけの殻だった。コービンとエフルッシのもう一人の助手のマダム・オーグがハエを管理していたが、彼はもうそれらのハエを実験に使うことはないだろうと感じていた10。ヨーロッパ中がそうであったように、フランスの遺伝学研究は停止に追い込まれていた。科学者達の多くは地下のレジスタンス運動の指導者になった。1940年の始めに、ビードルとマリオンはエフルッシのただ一人の子供だったイレーヌを安全のために引き取り一緒に住もうと提案した。しかしオルレアンは比較的安全だったから、エフルッシと妻のラジャはそこに残ることに決めた。エフルッシはビードル夫妻に次のように書き送った。 「それでも、私達はあなた方ご夫妻の提案を、将来も有効だと考えて心に留めておきます。このようなご提案を頂き、いずれにしても、お二人に感謝致します」 11。
1940年が終わる頃、フランスのドイツ占領地域に留まることはユダヤ人家族にとってますます危険だと感じたエフルッシは脱出を決意した。彼は、脱出に向けた情勢分析と経済的支援の可能性について、デメレックや他の人々と連絡を取った。1941年1月に、ナチス・ドイツの国家秘密警察組織ゲシュタポが彼と家族の逮捕を狙っているという警告を受けた。翌日早朝、荷物を何も持たず極秘裏にエフルッシの家族は父とルイ・ラプキン婦人とその娘を伴って脱出した12。彼らは、ユダヤ人だと悟られてはならないから、身便証明書のチェックがないことを祈りつつ夜行列車でパリからフランス南西部バスク地方の中心都市であるバイヨンヌへ向かった。バイヨンヌへ着くと、エフルッシの同僚の一人が引き合わせてくれたある男が彼らをドイツ占領地域から非占領地域への道案内をした。クレモント・フェランから彼らはリビエラのジュアン・ル・ピンへ向い、そこで不安な数ヶ月を過ごした後に、ルイ・ラプキンの手配で合衆国行きのビザを取得することができた。彼らはマルセイユからスペインのバルセロナを経てポルトガルのリスボンへ合法的な旅を続け、そこから蒸気船エクスカリバー号に乗船してニューヨークへ船で向かった(注:エクスカリバー号は、イギリス・アーサー王の伝説で登場する魔法の力が宿るとされる王の剣の名称をとって名付けられた)。コールド・スプリング・ハーバーのドブジャンシキーの家で夏の間しばらく逗留した後に、エフルッシの家族はバルチモアに到着した。ロックフェラー財団は彼にフェローシップを用意してジョンス・ホプキンス大学遺伝学科で1941年9月1日から始まる2年間の研究滞在を可能にした(注:ジョンス・ホプキンス大学はメリーランド州ボルチモアの私立大学)。エフルッシは1942年から1943学年度の遺伝学の准教授に任命された13。
エフルッシはジョンス・ホプキンス大学に3年留まり、その間ショウジョウバエの眼色素について遺伝学的な発達よりは化学的組成の他に性質と量の測定に焦点を当てた。ボルチモアでの経験は自由フランス軍に参加するためにロンドンへ向かうまでの 「引き延ばし作戦」 として役に立った。彼の自由フランス軍での仕事は連合国軍によるノルマンディー上陸作戦期間中の爆撃効果を評価することだった。戦争が終わって彼はパリへ戻りフランスの遺伝学分野で最初の教授に任命された。
エディンバラの国際遺伝学会議での総会講演で、ビードルはテータム、エフルッシとコービンとともにショウジョウバエの眼色の遺伝的制御について明らかにした研究の概要を語った。ビードルは講演14を、他の研究者の仕事のうち特にR.・スコット-モンクエイフ、J. B. S.・ホールデンとジョン・イネス園芸研究所の彼らの共同研究者達が行った園芸用植物の様々な花色を説明する植物色素の形成に関する仕事15の他に、アーチバルド E.・ギャロッドのアルカプトン尿症すなわちアミノ酸のフェニールアラニンとチロシンをホモゲンチジン酸を越えて代謝できずホモゲンチジン酸が体内に蓄積する先天的代謝異常の研究を引用することで始めた16(注:アルカプトン尿症の患者の尿は排出されると空気に触れて酸化し黒色を呈する。人で初めて見つかった劣性突然変異による先天的代謝異常である)。ビードルは 「これらのプロセスに関わる酵素システムの欠陥あるいは欠損はひとつの遺伝子の置換で生じる変化に帰すことができる」 と推論した。ビードルは彼が引用した研究対象に潜む遺伝子と酵素の関係に明らかに気がついてはいたが、そうした対象がその関係を追求するに十分に強固であるとはその時まだ考えてはいなかったようである17。
ビードルは、ショウジョウバエの眼色の発達について、 「この過程はそれぞれが遺伝子で制御される連続で平行なひとつの反応体系として進行する」 と結論した。講演の最後には、案全体は未だ暫定的でより多くの情報が入手可能となるとともに修正される可能性があることを認めた上で、案の本質的な正しさを指摘した。 「酵素の仲介を通じて作用する遺伝子」 がショウジョウバエの眼色素の形成に関与すると主張する彼とテータムの推論はまだ酵素システムに関する直接的な知識を欠いた 「純粋に希望的な憶測」 であることを彼は認めた。それでも 「どのような生物反応システムであれ、酵素こそが様々な段階の触媒として関与するに違いない」 ことを 「私達は知っている」 と付け加え、自分達の提案の合理性を弁護した。さらに 「蓄積されつつある証拠に基づけば、遺伝子の特異性は酵素の特異性とほぼ同じレベルであると確信する他ないことから、私達はこの推定が正しいと判断せざるを得ない」 と述べた。さらに推論を進めて、 「多くの遺伝子の直接的な産物は酵素か酵素を構成するタンパク質である」 と明確に述べた上で、 「現時点では、私達が自由な解釈に用いることのできる実験事実による限りは、残念だがこの推定に基づいて仮説を入念につくり上げることを正当化することはおそらくまだできない」 ことも付け加えて述べた18。ビードルの結論は、 「推論の重要性は、それが遺伝子の役割について多くを教えてくれるからではなく、むしろ遺伝学者と生化学者のどちらにもますます重要となると期待できる問題解決の方法を教えてくれるからである」 だった。ビードルは以前、次のような取組を提案したことがあった。 「遺伝学は様々なやり方で生化学に役立つし、遺伝学者が生化学者との連携から恩恵を得るもっと多くのやり方がきっとある」 19。生化学の正式な教育を受けていなかったが、それでもビードルは生化学と遺伝学のアプローチは互いに補完的であるという彼が後の多くの場面で繰り返し述べた見解が正しいと確信していた。おそらくv物質がトリプトファンの既知の代謝産物であることを同定できなかった教訓が、生化学の重要性を彼にはっきりと認識させたのだろう。
ビードルとエフルッシの長く続いた共同研究が生んだ成果は遺伝子が代謝経路の個々の段階を制御する事実を明確にした。v物質がトリプトファンの既知の代謝産物であることを同定できなかったことは、ショウジョウバエのシステムも現在どこかで研究されている他のどんなシステムも遺伝子作用のより広範な生理学的問題の探求、ビードルの言葉では生化学遺伝学の探求のための最適なシステムではないという信念をビードルに改めて確認させることになった。ビードルは突然変異が容易に認識できる既知の生理学的機能に影響するより適切な生物システムと実験戦略が必要であることを明確に理解した。もし解析の対象となる突然変異がショウジョウバエの眼色の場合にそうであったような未知の化学物質の生産ではなく、既知の化学物質の生産に影響するような遺伝子を対象とすれば、当該遺伝子の生理学的機能を調べることはずっと容易だろうと考えるのは論理的だった。
テータムとともに遺伝子と酵素の論理的な枠組みを実験的に追いかける最良の方法を見つけようと奮闘していたビードルは、テータムが1940-1941学年期の冬学期に担当した比較生化学のコースで行った講義のひとつを聞いてテータムが述べた何かに衝撃を受けた。ずっと後になってこの時のことを思い出してビードルは次のように語っている。 「テータムの講義のひとつを聞いたとき、彼が黒板に一連の生化学反応の順序を書いているとき、私は突然それまで何年もの間、自分達は何と愚かだったのかと気づかされた。有能な生化学者達が既に明らかにした酵素反応のすべてがそこにあった。もし遺伝子と酵素の概念が正しいとするなら、酵素が触媒する特異反応に関与する遺伝子を即座に同定できる。だから、やり方を逆にすればいい。既知の遺伝子が制御する酵素反応を探すのではなく、既に知られている化学反応を制御する遺伝子を探すことで厄介な化学の問題を避けることができる。では、どうするか?答えは簡単だ。たくさんの既に知られた最終産物の生産にかかわる一連の反応が現に行われている生物をまず選ぶ。次に、一連の反応の特異的な段階をそれぞれ制御する遺伝子を探せばよい」 。ビードルの論理は実に明確で単純で洗練されたものだった。しかしどんなアイディアも明確な実験手法に帰着されなければならない。 「実際上、生物の全ての反応は生化学反応であるから個々の反応は連続した特異反応に分解される筈である。もし細胞における全てのそうした合成反応が遺伝的であるなら、合成反応を制御する遺伝子に突然変異を誘導することで特異物質の生産をブロックすることが可能である。私達は次に突然変異を探すことになるだろう。そのような突然変異はビタミン、アミノ酸やその他の細胞に必須な基本構成要素であり、本質的に細胞質内で拡散可能な物質の合成能力を失った変異体として選抜できるはずだ」 20。ビードルはこの手法が成功することを確信した。
テータムの講義の一体何がビードルのひらめきの引き金になったのだろう。幸いにも講義の内容が医学細菌学科の若手教員だったチャールトン・シュワールトの手書きのノートに残されていた21。1941年1月から2月の初めまで、テータムは講義のなかで微生物、植物と動物の中間代謝について当時知られていた事実に関する包括的なレビューを行った。テータムは微生物のもつビタミン、アミノ酸、プリン塩基、ピリミジン塩基などに関する多種多様な栄養要求性は、それらの微生物がより単純な前駆物質からこれらの代謝産物を合成できない事実の反映であると主張した。より明確に言えば、微生物の示す複雑な栄養要求性はある必要な栄養素の生産に不可欠なひとつかそれ以上の遺伝子の機能が失われていることの反映であるとする既に他の研究者が発見していた見解を、彼は改めて総括し主張したのだった22。
講義の最後にテータムは重要な疑問をさらに誇張して、 「遺伝子は一体何をするのだろう?」 と述べた。彼は、代謝反応を制御する機能を持つのは遺伝子で、さらに進化の過程で蓄積した突然変異がある特定の代謝機能の喪失をそれぞれの種にもたらしたと結論した。彼は微生物がアミノ酸、ビタミンなどの合成能力の点で互いに異なっている多くの例を示し、ヒトについても様々なアミノ酸やビタミンの合成ができない理由はそれらに関わる反応が対応する遺伝子の機能喪失によりおそらく抑止されているからだと推論した。しかしテータムは、遺伝子がどのように代謝反応を制御するのかという点については、遺伝子が果たす決定的な役割を洞察することには失敗していた。
ビードルは、もし既知の栄養素の欠損を引き起こす突然変異が見つかれば、それらを解析することで遺伝子の作用と代謝過程を直接結びつけることができることを即座に理解した。しかしこれをうまくやるには、できるだけ数は少ないが明確な栄養要求性を示す生物種が必要だった。そうした生物種は同時に放射線やその他の突然変異源による突然変異の誘発が可能でなければならない。さらに、そうした生物種は速やかな遺伝解析が可能で、それによってどんな栄養要求性もひとつの遺伝子、可能であれば特定染色体上にマップできるひとつの特定遺伝子に帰属させうるようなものが望ましい。テータムはこうした必要に合致した微生物種を近くの医学細菌学科に尋ねてみたが、有益な助言を得ることができなかった23。候補微生物のひとつは子嚢菌のアカパンカビ属(Neurospora)で、それはビードルが既にそれらの性質と遺伝学を熟知していたカビの一種だった。
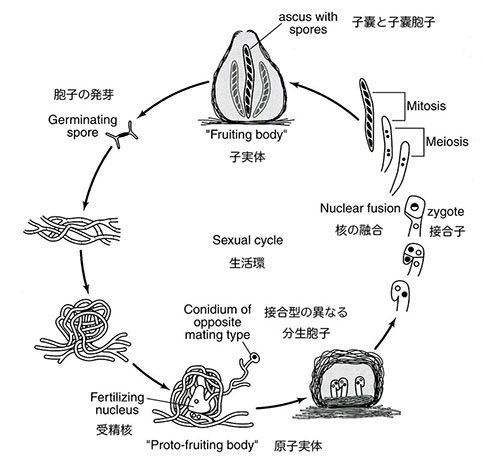
アカパンカビはジャワ島を旅行中だった有名なオランダ人植物学者フリードリッヒ A. F. C.・ウェントの注意を引き、彼によって多くの異なる種が記載され登録されていた。どこにでもいるカビで、その最も際立った特徴は様々な栄養繁殖体の表面が鮮やかなサーモン・オレンジ色の 「産毛」 で覆われていることである。アカパンカビ属が登場し遺伝学の研究材料となったのは1927年のことだった(注:アカパンカビは酵母やカビを含む子嚢菌門に属する糸状菌)。このカビがビードルを引きつけた理由は、それが無性的に半数体として繁殖可能であることと同時に生殖サイクルを人為的に誘導できることだった。ショウジョウバエと同様に、アカパンカビにはふたつの交配型が存在し、それらの間で受精が起こると減数分裂時の交叉産物が生じた胞子中に保存される(注:減数分裂によって作られる胞子を子嚢胞子と呼び、子嚢胞子を内部に保存する袋状の構造を胞子嚢と呼ぶ)。さらに、胞子中の遺伝マーカーの分離パターンが染色体上の遺伝子の並びを決定するための実用的な方法を提供する。しかしビードルの目的にとってこのカビがもつ生殖様式の最も重要な点は、栄養要求性の変化を引き起こす突然変異が特定の単一遺伝子の変化であることが確認できること、さらに特定染色体上に遺伝子の位置を決定できることだった。
ビードルがこのカビについて初めて話しを聞いたのはコーネルの大学院生の時で、ニューヨーク植物園の有名な植物病理学者バーナード O.・ドッジがカビの細胞遺伝学について講演をした時のことだった24。ドッジは、カビの生活環と配偶行動のほとんど全て、特に胞子の形成に至る減数分裂の詳細を明らかにすることでカビ遺伝学の基礎を作った科学者である(注:生活環は栄養生長世代と配偶子世代が交代する核相交代を含む生殖による世代交代の意)。彼の幸運ともいえる発見、すなわち生活環中の生殖相で作られる胞子が保存できること、温めることで発芽が起こることはこのカビの遺伝学研究を飛躍的に発展させた。
ドッジは、ウイスコンシンの父の農場を助ける必要から、高等学校を卒業したのは二十歳になった時だった。彼は高等学校で15年以上、教師生活を続けながら断続的にウイスコンシン大学で学び、その後にコロンビア大学で糸状菌学の学位を取得した。ドッジがカビで注目すべき研究を成し遂げたのは、ニューヨーク植物園の植物病理学主任となった56才の時だった25。ドッジはカビがショウジョウバエとともに遺伝学で脚光を浴びるに相応しい材料をなることを確信し、近しい友人だったモルガンにカビの培養物をパサディナへ持って行くよう説得した。しかし新しい研究室でカビの仕事を始めようとしたモルガンの試みは、残念ながらショウジョウバエの培地にしばしば生える細菌と普通のパンカビであるペニシリウム(アオカビ)による汚染が原因で失敗に終わった。それでもモルガンは、カール C.・リンデグレンがPh.D.コースの学生としてカルテックの新しい生物学部門に入学すると、彼にカビを研究対象とするよう提案した。スターテバントとブリッジスの大きな支えと熱心な助言によって、リンデグレンはカビの基礎遺伝学を前に進め、新しい遺伝子を発見し、遺伝的な交配子孫の検定法を開発し、染色体地図の作成に向けた最初の一歩を踏み出すことができた26。ビードルが1931年にカルテックに着任した時にはリンデグレンはまだカビの研究をしており、このカビの潜在的な有用性をよく理解していたビードルはスターテバンとともに教科書 「遺伝学入門」 にカビの遺伝学に関するレビューを付け加えた27。
テータムが講義で遺伝子機能に関する疑問や課題を提示した頃、ビードルはドッジに手紙を書いて、 「テータム博士と私はカビの栄養に関する仕事に、最終的にはカビの栄養要求性が遺伝的な仕組みに依存するか否かを決める仕事に興味を抱いています」 と伝えた。ビードルはドッジに 「もし予備的な実験の結果が有望なら、私達は入手可能なカビの各種と様々な野生型を試験してみたいと希望しています」 と伝えた28。2週間後には、テータムが行った異なる起源をもつ3系統の調査から、新しく発見されたビタミンであるビオチンの添加なしにはどの系統も生育しないことが明らかとなり、ビードルはこの事実をドッジに報告した29。ビオチンがユトレヒト大学のF.・クーグル教授のグループによって分離され酵母と糸状菌の生育に必要な成長物質として同定された時にたまたまクーグル教授のもとに滞在していたことから、テータムはビオチンについてよく理解していた30。偶然だが、ユトレヒトでテータムと実験室を共有していたニルス・フリースが広範な糸状菌の種を対象にビオチン要求性を調査していた。その過程でフリースはカビを育てる単純な餌を開発した。このカビ用培地には糖(何種類かの糖類が有効)、窒素源(アンモニアか硝酸塩)、リン(リン酸塩の形で)といくつかの塩類とビオチンが含まれていた。フリースと呼ばれるこの培地はアカパンカビ属の研究で今でも用いられている。当時ビオチンはまれにしか手に入らない化合物だったが、過去からのクーグル実験室とのつながりから、テータムはすぐにビオチンを入手できた31。
|
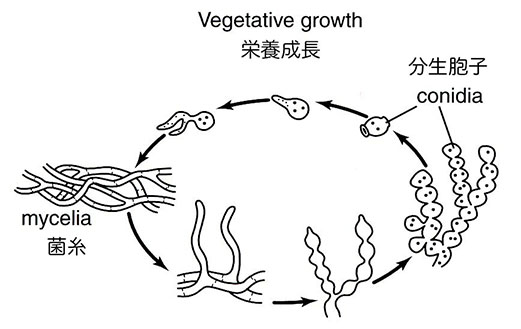
カビの生育行動とその単純な遺伝的性質は、速やかに同定可能で特定の栄養素を欠乏した突然変異体の獲得を目指したビードルとテータムの計画にとって重要な必要条件だった。全ての必須栄養素を含む固形培地の表面で、彼らが最終的に選択したアカパンカビ(Neurospora crassa)は無性的に増殖し、菌糸体あるいは菌糸と呼ばれる高度に枝分かれした管状の繊維の絡まったマットを形成する。隔壁が菌糸を大きさの異なる節に分け、その内部に核が他の細胞質内器官とともに分布する。生育は菌糸の先端部が伸長することで起こり、先端部の伸長速度がカビの生育の目安となるが、これはビードルが有効利用したこのカビの特徴的な性質のひとつだった。時折、特有の遊離菌糸あるいは表面菌糸が分生胞子と呼ばれる球状でサーモンピンク色をした構造を分化するが、分生胞子は培養物に軽い気流を与えることで容易に培地表面に分散する。適当な培養条件下では、分散した分生胞子は発芽して新たな菌糸の生育を開始する。栄養繁殖期と呼ばれるこの生育段階では菌糸と分生胞子の核は半数体であり、アカパンカビの7本の染色体のそれぞれ一組をもつ。
|
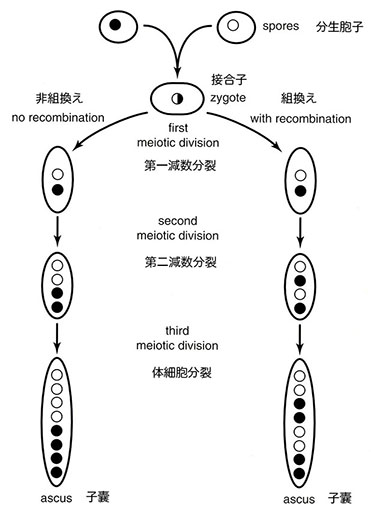
アカパンカビは人為的に栄養繁殖相から生殖相へ移すことができることから、ビードルには極めて魅力的な材料だった。半数体細胞はA型あるいはa型と呼ばれるふたつの交配型のどちらかとして存在する。飢餓状態に置かれると、菌糸体の一部が 「子実体」 に分化して、その内部で半数核のA型あるいはa型の分生胞子が融合しふたつの型の核をもつ細胞が作られる。核が融合すると細胞は伸長して嚢状の子嚢と呼ばれる構造物を作り、その内部で2倍体核がふたつの連続した細胞分裂(減数分裂)を行う。この過程で4つの半数体胞子が形成され、それぞれがさらに一回の体細胞分裂を行って8個すなわち4対の子嚢胞子ができる。この時点で、それぞれの子嚢胞子の周囲に細胞壁が形成される。
減数分裂の第一分裂で作られた二つの細胞は横に並び、第二分裂で生じた4つの細胞はそれぞれが横並びの対として配置される。4つの細胞のそれぞれは次に一回の体細胞分裂を行い8個の子嚢胞子ができる。結果として、上部の4つの子嚢胞子は第一分裂で分かれた二つの細胞のひとつに由来し、下部の4つの子嚢胞子は第一分裂で生じたもうひとつの細胞に由来することになる。重要な点は、連続した分裂によりできた子嚢胞子が子嚢のなかで特徴的な空間配置を占めること、特に4対の子嚢胞子が直線的な空間配置をとることである(注:8の子嚢胞子を上下4つずつに別ける面が減数第一分裂の分裂面、上下4つをふたつずつに別ける面が減数第二分裂面である)。アカパンカビの遺伝学者は解剖針を使って個々の子嚢を押し開けて内部から子嚢胞子を順番にひとつずつ取り出すことで、順序を壊さずに並んだ子嚢胞子を子嚢中で形成され配置された順番どおりに分析するができる。
|
各子嚢中のそれぞれ2個ずつ4対の子嚢胞子の遺伝解析により、生殖過程を始めた二つの半数体分生胞子の遺伝子型とともに減数分裂で起こった事象の推定が可能となる。例えば、A型とa型の交配型の違いは子嚢胞子がひとつの交配型決定遺伝子のどちらのアリルをもつかで決まるが、分裂後の子嚢中には4つのA型アリルをもつ子嚢胞子と4つのa型子アリルをもつ子嚢胞子ができる。同様に、野生型と栄養要求性突然変異体の交配では、4つの子嚢胞子は野生型アリルを残りの4つは突然変異型アリルをもつことになる。もしひとつ以上の突然変異が表現型を決める場合には、これとは明瞭に異なる子嚢胞子の種類と配置が観察される。
加えて、子嚢中の8個の子嚢胞子の配置は減数分裂で組換えが起こったか否か、さらに組換えの頻度を反映する点で重要な情報を与える。もし組換えが起こらなければ、子嚢胞子は一方のアリルをもつ4つの子嚢胞子が他方の型の4つの子嚢胞子と隣接して子嚢中に並ぶ(4-4配置)ことになる。他方、もし減数分裂で組換えが起これば、子嚢胞子は対の状態すなわち一方の型の一組が他方の型の一組みと隣接して並ぶことになる(2-2配置)(注:ここで検出される組換えは、問題とする遺伝子と動原体との間に生じた組換えである。遺伝子より染色体の末端側で生じた組換えは検出できない)。2-2配置を示す子嚢胞子を含む子嚢の数の4-4配置を示す子嚢胞子を含む全子嚢の数に対する相対頻度で減数分裂時の組換え頻度が測定できる。このように子嚢中の4組の子嚢胞子の遺伝子型と表現型によって、二つの交配型が単一の遺伝子の違いによるか否かを決定できる。例えば、ビードルは栄養素を欠損した分離株がひとつの突然変異に由来することを野生型との交配で生じる子嚢胞子の配置パターンから決定することができた。もしそのような交配で4つの突然変異型を示す子嚢胞子と4つの野生型の子嚢胞子が得られれば、栄養要求性は単一の突然変異の結果であると判断できる。同一の栄養要求性を示す独立に分離した系統が野生型との交配で4-4配置と2-2配置を同一頻度で生じれば、それらは同一遺伝子のアリルであると結論できる。一方、同一の栄養要求性を示す独立に分離した系統が異なる子嚢中でこの2種類の子嚢胞子の配置と明確に異なる相対頻度を示したとすれば、二つの突然変異は異なる遺伝子に起因すると結論できる。
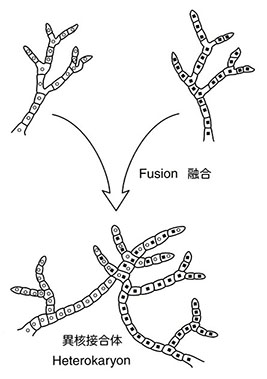
アカパンカビの示すもうひとつの有用な性質は同じ交配型の菌糸が融合する能力をもつ事実である。この場合には二つの半数体核は細胞質中で融合せずに残る。もし生じた菌糸が異なる遺伝子型をもつ核を含むなら、そのような菌糸を構成する細胞はヘテロカリオンと呼ばれる。遺伝的な意味では、ヘテロカリオンは同じ核中に二つの異なる半数ゲノムをもつ二倍体の動物や植物の細胞と同等である。もし野生型と突然変異型の菌糸から作ったヘテロカリオンが野生型の形質を示せば、突然変異は劣性であると判定できる。一方、ヘテロカリオンが突然変異型ならば突然変異は優性である。異なる遺伝子に由来する突然変異をもつ系統間で作ったヘテロカリオンは、それぞれの半数体核が他方で失われている野生型アリルをもつことから野生型の形質を示す。一方、突然変異が同一の遺伝子に起こった系統間で作ったヘテロカリオンでは、どちらの系統も正常なアリルをもたないから突然変異型の表現型が現れる(注:一般に、同一表現型と判定される起源の異なる突然変異が同一の遺伝子によるか別の遺伝子によるかを決めるには、それらの雑種を作り雑種の表現型を見る相補性検定と呼ばれる検定法が有効である。相補性検定は一般に以下のやり方で行われる。二つの突然変異体を交配し得られた雑種の表現型を調べる。もし雑種が野生型を示せば二つの突然変異は相補的である、すなわち異なる遺伝子に由来すると判定する。一方、そのような雑種が同一の突然変異型を示す場合には、二つの突然変異は相補的でない、すなわち同一の遺伝子に由来すると判定できる)。
|
従って、ビードルとテータムは突然変異体を調べる二つの方法、すなわち突然変異体を野生型と交配して子嚢胞子の配置を調べる方法と突然変異体と野生型の間あるいは二つの突然変異体間で作成したヘテロカイロンの表現型を見る方法で検定することができた。こうして彼らは、同一の栄養要求性を示す独立に得た突然変異体が同一遺伝子の変異によるのか異なる遺伝子の変異によるのかを容易に決定することができた。彼らには突然変異を特定の染色体上に位置づけることも可能だった。
このようにアカパンカビは、それぞれの突然変異が単純なフリース培地で生育する能力を回復するためにはどんな栄養素が必要か決定することで同定可能な栄養要求性をもつ突然変異体を発見し、それを性質づけるために完全に適した材料だった。ビードルとテータムはこの戦略の有効性を確信したが、それはショウジョウバエの眼色に影響を与える単一の突然変異が単一の反応を抑止するという彼らの信念によって支えられていた。しかし多くの遺伝学者は、こうした可能性をもつ実験に気がついてはいても懐疑的であったが、それはひとつの遺伝子とひとつの生化学反応の間に直接的な関係があるとは思えないというだけの理由からだったのだろう。この懐疑は、どのような形質あるいは生化学的機能であろうと、それらの決定には複数の遺伝子が必要であるとする考え方から予想できたことだった。個々の遺伝子が複数の機能をもつことを疑う理由はありそうもなかったし、突然変異はより重大な結果、おそらく複数機能の欠損を引き起こすであろうと考えられていたからである。マラーは10年以上も前に既にこの見解を述べていた。マラーによれば、 「例えば、ある遺伝子の違いが特定の酵素の存在あるいは欠損の原因だと分かったとしても、その遺伝子が直接に酵素を生産したことの証明にはならない。酵素の生産は一連の中間過程を経て生じたのかもしれない。すなわち、その酵素を不活化するか破壊する酸が生産されるとする、その酸は実は他の酵素で生産され、その酵素は補酵素で活性化され、その補酵素はあるタンパク質で作られる、しかもそのタンパク質は遺伝子によってイオン化される。例えれば、この家を建てたのは誰だと確定的に言うことができないのと同じではないだろうか?」 32。実験操作では遺伝子の作用を理解することはできないという考えから、もっと悲観的な者も多かった。実際の実験が始まって5年が過ぎた時でも、マラーは遺伝子と酵素の関係の実体について態度を曖昧にした。しかしビードルとテータムは自分達が正しい道に立っていると信じて疑わなかった。彼らの唯一の気がかりは、突然変異の頻度が低すぎて求める突然変異体が容易に得られないことだった。
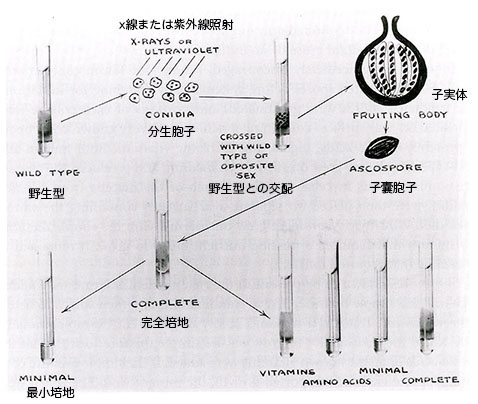
彼らはアカパンカビの分生胞子に様々な強度のX線を照射することで突然変異の誘発を試みた。物理学科が自製のX線照射装置を作ってくれたが、それは 「放射線をまき散らす代物」 だったから、装置の周りは金槌で叩いて作った鉛の板で囲む必要があった33。紫外線を分生胞子に照射しても突然変異が得られた。彼らは個々の突然変異を独立なひとつの事象としてとらえる目的で、照射した個々の培養からひとつずつ分生胞子を選んで遺伝実験に供した34。
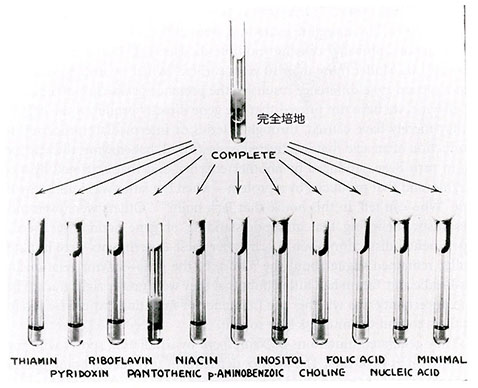
もちろん始めから失望を味わいたくはなかったので、ビードルとテータムは突然変異の探索に本格的に取り組む前に、まず5,000個の分生胞子を集めて保存した。X線や紫外線を照射した分生胞子を発芽させて、それぞれの培養の一部をタンパク質、核酸、糖の既知の全構成要素と全てのビタミンの混合物を含む寒天で固化した完全培地の表面に塗布した。こうした栄養価の高い完全培地上では突然変異体と野生型のどちらも生育可能で特徴的な繊維状の菌糸体を作る。完全培地上の個々の菌糸体から得たサンプルを次に単純なフリース培地を含む個々の寒天培地上に移植する。野生型のアカパンカビはフリースの最小培地で正常に育つが、必要なある栄養素が欠損した突然変異体は育たない。従って、次にやるべきことは失われた栄養素を決めることだった。
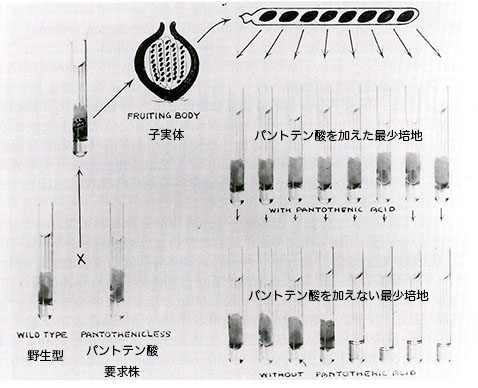
そこで、完全培地に含まれることが分かっている栄養物質について、始めに各栄養要素を組み合せ次にはそれらを個々に最小培地に加えてみてどれが突然変異体の生育能力を回復できるかを決める。例えば、突然変異体を全てのアミノ酸あるいは全てのビタミンを含む最小培地に植えて生育能力を試験する。もし全てのビタミンを加えた最小培地で生育できれば、次は突然変異体をビタミンのグループごとに試験し、もし生育が観察できればそのグループのビタミンを個々に試験する。こうして各突然変異体が必要とする特定のビタミン、アミノ酸あるいはそれらの前駆物質でさえも個別に決定することができた。完全培地では育つがどの既知の栄養素を加えた最小培地でも育たなかったとすれば、突然変異体は何か未知の必須要素を合成できないと考えられる。未知の栄養素は完全培地に含まれるのだから、それを分離し同定することが可能である。突然変異体の候補は、次に異なる交配型の野生型と交配して単一遺伝子の違いによるかどうかを決定する。もし突然変異の候補と交配型の異なる野生型の交配から最小培地で育つ子嚢胞子4つと育たない子嚢胞子4つが得られれば、突然変異は単一遺伝子の変化によると判断できる。
ビードルとテータムは299個の子嚢胞子を調べて最初の突然変異体を見いだすことができた。アカパンカビの培養を入手し生育に必要な栄養素の推定を始めて3ヶ月以内に、ビードルはリンデグレンに 「アカパンカビは幸運をくれた」 と伝え、さらに 「このカビは仕事のやり易い素敵な奴だが、その有利さはほとんどまだ評価されていない」 と付け加えた35。2週間後には同じニュースがドッジに届いた36。それから二ヶ月の内に、ビードルとテータムは方法論の詳細を含めて3つの栄養要求性突然変異体を得た事実を記述した論文を発表した37。ひとつの突然変異体はビタミンB6(ピリドキシン)を、2番目はパラアミノ安息香酸(葉酸の構成要素)を、3番目はビタミンB1(チアミン)を要求する突然変異体だった。ビタミンB1がチアゾールとピリミジンという二つの構成部分からできていることを二人は知っており、当該の突然変異体はピリミジン部分を合成できないことも決めることができた。論文の提出は1941年10月8日だったが、合衆国科学アカデミー紀要の11月号に掲載されたことから判断すれば、この論文は科学上の 「爆弾」 と評価されて緊急出版の取扱いを受けたに違いなかった。
ビードルは、ショウジョウバエの眼色の仕事で定義されたやり方を用いれば、新しい発見が遺伝学と生化学の統合を可能とすることを理解していた。アカパンカビのもつ類いない特性が形態的な性質だけでなく代謝の遺伝的制御を探求するための実験上の楔となって突破口を開いた。換言すれば、遺伝子作用の仕組みが研究の手の届く範囲に入ったのだった。この目標をより精力的に追求するには、研究グループを格段に大きくする必要と特に生化学の訓練を受けた研究者を増やす必要があった。カルテックは優れた若手研究者を勧誘する最適の場であると思われたので、論文が投稿されるとすぐにビードルはスタンフォードで彼のグループに加わる研究者を捜し始めた。
スタンフォードでのビードルのセミナー発表会に参加した研究者達はその発表内容に圧倒された。どんなタイトルを思いついたところで、セミナーで何が始まるかを事前に知らせるに相応しいものなどなかっただろう。参加者の誰もが予想していたように、セミナーはケルクホッフで行われるいつもの一般生物学セミナーのひとつの筈だった。セミナーの鮮明な印象を記録したノートのひとつには以下のような記述がある。 「ビードルの話しはたった30分で終了した。話しが突然終わったとき、セミナー室は沈黙で満たされた。だが、沈黙は賛辞の表明だった。そのような発見をした研究者がたった30分で話しをやめるとは私達聴衆には考えもつかないことだった。それにも増して誰もが何か歴史的なことが起こったのだと理解した。私達は皆、たった今起こった革命的な発見が次に何をもたらすだろうかと可能なかぎり思案を巡らせていたのだと思う。ついにビードルが実際に話し終えたことに気がつくと、フリッツ・ウェント教授が立ち上がり、実は彼の父親こそアカパンカビを用いて最初の研究を行った研究者だったが、セミナー室の大学院生達に明らかに熱狂した様子で告げた。ビードル教授の講義は生物学が既に終わった学問ではないこと、まだまだ偉大な発見が行われる対象であることを見事に証明した!」 38。いつもの様に質問がいくつかあったが、ほとんどはビードルが予想していたものだった。いくつかには答えが用意されていたし、いくつかには実験中であると答えた。セミナーは科学的にも演出の上でもまさに 「力作」 だった。この講演以外のビードルの講演については、既に何度か本書で語っているから、読者の皆様にはきっとビードルの過去の発表に対するモルガンの批判がどれだけビードルの発表スタイルの改善に役立ったか明らかだろう。もちろんビードルの電撃的な発表内容それ自体が間違いなく発表スタイルの欠点を遥かに凌いだものだった。
ビードルとテータムの発見の報が伝わるにつれて、スタンフォード医学部の学部1年生や2年生までもがその意味に興奮を覚えた39。この時までジョンス・ホプキンス大学でショウジョウバエの眼色の仕事を続けていたエフルッシは、いつもの大げさな書きぶりでビードルに賞賛の意を伝えた。 「貴兄とテータムさんにおめでとうと言わせてください。最初のこうした結果から、私は間違いなく貴兄達が最も有望な可能性をもつ未だ探求されていない領域に入りつつあるのだと確信します」 40。エフルッシは、二人の長い交流から、ショウジョウバエの眼色の仕事に関する共同研究について期待していると伝えたが、ビードルの返事はもっぱら最近新しく獲得したアカパンカビの栄養要求性突然変異体について語っていた。ハエ系統に関するいくつかのデータと処置に関する質問に対するビードルの答えは、 「ショウジョウバエに興味をもつ学生が見つけられなければ、ショウジョウバエに戻ることはないと思います。アカパンカビの仕事は常に加速の最中で、私はますますこれにのめり込んでいます。この状態はハエ実験室で仕事をする誰にも共通で、ショウジョウバエはスタンフォードでは少なくとも一時的に運のない研究対象になったようです」 だった41。ビードルは二度とショウジョウバエに戻ることがなかったが、エフリッシはその後暫く眼色素の研究を続け、結局その後に他の領域へ移って行った。
当時、ビードルの研究はロックフェラー財団からスタンフォードへの10年間計20万ドルの補助金で支援されていた。しかし、これらの資金のうち彼の持ち分は増え続ける活動ペースに加えて補充しなければならない研究者を支えるには不十分だった。革新的な研究の支援に対して限定的ではあるが歴史的に奨学資金を提供してきたことで知られるアメリカ哲学協会のペンローズ基金から支給されるビードルとテータムの初期研究への支援金は僅かずつ増加してはいたが、彼らへの研究資金提供を実質的に増やすことができるのはロックフェラー財団のみだった。最初の論文が出版された2週間以内にビードルはロックフェラー財団のウィーバーに手紙を書き、大きな突破口が開かれたことを伝えるともに、彼らの戦略を支える根本方針と方法の詳細を伝えた。それまでに、ビードルとテータムは一ダースを越える確かな突然変異体を蓄積していた。ビードルは、 「私達には生化学遺伝学にとって有益なだけでなく大きな潜在的価値をもつ方法論がある」 と意義を強調した。彼には単純で信頼性の高いビタミンとアミノ酸の検定法が仕事の副産物として手に入るだろうとの見通しがあった42。ビードルは既存の方法論より特異的で速やかにビタミンとアミノ酸を検定できる突然変異体の有利さを強調したが、それは今後も続くことになるロックフェラー財団への要請の始まりだった。
ロックフェラー財団が研究支援金を増額してくれると期待したビードルは、1941年12月中頃にニューヨークでその可能性を議論したいとウィーバーに申し出た43。実は一週間前に日本によるパール・ハーバ攻撃があり、引き続いて合衆国の宣戦布告があったばかりだったが、ビードルは、その性質からして、これを理由にウィーバーとの会見交渉計画の実施に躊躇するようなことはなかった。日本軍の侵攻の可能性というような実際上の暗転がなければ、ニューヨークへ行くとビードルは決めていた。合衆国が戦争に突入して数ヶ月以内のビードルとロックフェラー財団との交渉から見ると、どちらにも優先度を変えた兆候は見られなかった。それでも、二人には戦争への取組みが生命科学の基礎研究に一定の制限を与えるだろうと予想ができた。ビードルは、研究を軌道に乗せておくためには基礎と応用をうまくやりくりする方法を見つけ必要があると理解した上で、新しい研究を前進させると心を決めた。
ウィーバーと会う前に、ビードル・グループの発見は価値ありと見たメルク社から、発見の全てに排他的な特許権を譲渡するという条件で、研究計画全体を対象に資金提供する用意があるとの意見表明があった(注:メルクは合衆国ニュージャージー州を拠点とする世界的な製薬企業。ドイツのダルムシュタットに本社を置くメルクとは起源は同一だが別の企業である)。メルクは、アカパンカビの仕事は新しい成長因子の同定に向けた彼ら自身の仕事を促進するだけでなく、ビードルに天然物の分離に必要な素材の他に高度な専門知識と技術を提供することで仕事の効率を上げることができると確信した。メルクにはビードルの計画遂行に必要なだけの十分な資金提供の用意があったが、ビードルはメルクとの共同研究契約に入る妥当性についてまずウィーバーに意見を求めた。スタンフォードのウィルバー学長と学科長のテイラーはメルクとのつながりを開拓するようビードルに勧めた。しかしビードルは、そのような取り決めが研究を促進する一方で製造権と特許権に関する問題を招致することになるのではと危惧した。彼は、スタンフォード大学の方針で、職員が研究で特許を求めることに反対である点もよく承知していた。メルクの職員からは、それならば特許権は除外してメルクの研究者に研究上の発見の排他的な 「第一目通し」 の権利を与える、そうした共同研究条件ではどうかと打診があった。
ロックフェラー職員とのミーティングの間、ビードルはハンソンに、 「自分は特許にも個人的な利益にも興味はないが、この仕事を前に進めるために外部資金が必要だ」 と訴えた。彼は、メルクの提案とは別に、アメリカ研究協力法人(RCA)から研究計画に対して10,000ドルの資金提供の申し出があったこともハンソンに伝えた。RCAは大学が得た特許権を譲り受け、その特許権使用料を使って有望な研究を支援するための非営利組織であった。しかしビードルは、これらの提案のどちらにも紛糾の種がありそうで心が進まないとハンセンに告白した。彼らとのミーティングの報告を読んだハンソンは、ビードルの第一希望は 「懸命に仕事し研究結果を自由に公表すること以外に何の義務もないロックフェラー財団からの資金である」 と察知した。ビードルの第2の選択はRCAで3番目がメルクだった。」 44。
ハンソンは長い間ビードルを 「生物学で同年代のうち最も有望なひとり」 と看做していた45。彼はアカパンカビの発見がもつ重大な意味をよく理解しており、ビードルの年間支援の増額に賛成だった。その時点でビードルは生物学科へ支給されていたロックフェラー財団の資金から年20,000ドルを得ていた。ハンソンは、財団は1942年に7,500ドルの追加資金を、その後の2年間は毎年度同額を提供するだろうとビードルに伝えた46。ビードルは、必要資金を定期的に査定してもらうために、資金は年度ごとに評価して欲しいと申し出た。グループの誰かを引き抜きで失えば資金の必要額が減る可能性があると気遣ったからだった。RCAからの資金提供はそれまでなかったが、ロックフェラー財団からの資金提供が不十分になればRCAが資金を提供するという約束もビードルは再確認した47。
パサディナでのセミナー講演を終え、研究資金を手にして、ビードルはカルテックでPh.D.を取得したばかりの二人、ノルマン H.・ホロウィッツとデビッド M.・ボナーをスタンフォードに招いてアカパンカビのプロジェクトに加えた。何年も前にビードルは、ホロウィッツのカルテック大学院への進学のために提出された申請書を検討した折りに、彼が既にピッツバーグ大学の学部学生として移植に関する論文を発表していたこと知り、審査評に 「褒め言葉」 を書き記していた。ピッツバーグ生まれのホロウィッツは遺伝学でPh.D.を取得するために1936年秋にカルテックにやってきていたが、モルガンは彼の研究テーマとしてはアルバート・タイラーとともに海生生物を対象にした発生学の仕事をするよう割り当てた。ホロウィッツは学位論文研究で二つの海生無脊椎動物の卵の形態的発達に伴う呼吸の特性と時間的な側面を調べた。彼は1939年に学位を取得し、その後に全米研究評議会(NRC)のフェローシップを得てヨーロッパでの海外研究の機会を得た。しかしヨーロッパにおける戦争で計画変更を余儀なくされた彼は、スタンフォードでダグラス・ウィタカー教授と発生学の研究を継続した。スタンフォードで彼は、そのときショウジョウバエの眼色の研究の最終段階に没頭していたビードルと再開し、関係を新たにした。その年の終わりに、仕事上の必要もあってホロウィッツはヘンリー・ボルスーク教授に請われてポストドクとしてカルテックへ戻り、地域の歯科医の資金提供を受けた歯形成の発生学に関するプロジェクトに従事した。その後ほどなくアカパンカビを対象にしたビードルの初期の発見に関する刺激的な講義を聴いたホロウィッツは、ビードルからスタンフォードでポストドクとして働くよう要請を受けて感激に震えるほどだった。少しの躊躇もなく申し出を受けたが、スタンフォード到着は1942年の半ばまで遅れた。ホロウィッツと家族は、大学の実験室までゆっくり自転車で10分、早足で歩けばそれぐらいの距離にあるキャンパスの角に家を借りた。戦時下のガソリン制限もあって車は贅沢品だった。
デビッド・ボナーがビードルのセミナー講演を聴いたのもPh.D.研究を修了したばかりの時で、彼もまたアカパンカビ・プロジェクトへの研究参加要請に熱心に応じた。ボナーはソルトレーク・シティーの大きなモルモン教一家で育ち、父は大学の化学の教授だった。彼は1937年に化学を専攻して学部を卒業した後、4人兄弟の最年長の兄ジェームスが生物学部門の教員を務めていたカルテックにやってきた。その当時、植物生理学のホットな話題は、どのようにして植物生長因子であるオーキシンが根、芽、花など植物の一定の部位のみで生長を促すのかであった。初めのうちはフリッツ H.・ウェントと、次にアリー・ハーゲンスミットと共同で仕事をしたボナーは、葉の成長に影響を与える生長因子を同定した。その後、彼は葉のオーキシンと他の植物組織に影響するオーキシンの関係を調査した。ビードルは、化学の経験と代謝への興味を持っていたボナーは個々のアカパンカビ突然変異体が引き起こす代謝の欠損を調べるのにうってつけだと考えた。ボナーとホロウィッツは同じ時期にスタンフォードに到着し、テータムの実験室に接した地下の一角で仕事を始めることになった。
ハーシェル K.・ミッチェルも、アカパンカビの実験を聞いてスタンフォードに心を向け、ビードルに自分を雇用してくれるよう熱心に懇願した一人だった。カリフォルニア生まれで、ポモナ・カレッジで化学を勉強したミッチェルは、この分野で既に二つの注目すべき成功を収めていた。オレゴン大学の修士コースの大学院生だったときに彼はビタミンのパントテン酸の構造を明らかにし、それから数年後には、テキサス大学のロジャー J.・ウイリアムスとともにPh.D.論文の一部として、葉酸の構造を明らかにした。ミッチェルのための奨学金はまだビードルの手元になかったが、ミッチェルの酵素に関する経験はビードルの眼に留まった。ビードルは自分の決定を後悔することが決してなかったが、ミッチェルの件でもそうだった。夫がサンフランシスコで職を得たばかりのマリー・ホウラハンは、もう一人の 「押し掛け」 だった。彼女は放射線生物学の仕事についてコールド・スプリング・ハーバーから得た強い推薦状を携えてきた。彼女はすばらしい仕事人で彼女を雇って後悔する者は決してないだろうと推薦状に書かれた確約に動かされて、ビードルは彼女をチームに加えた。何年か後に、マリーとミッチェルはそれぞれ連れ合いと離婚して結婚することになった。
比較的よく訓練されたポスドクの他に新しい研究事業への参加を望む多くの大学院生があった。もちろんビードルには彼らをサポートする資金を見つける必要があった。幸いなことに、明確に決定された特定の栄養素を欠いたアカパンカビの突然変異体は生物検定に用いることができるという期待から、栄養機構がロックフェラー財団の支援でこの方向に沿った仕事をする約束で、何人かの大学院生のための奨学金の支給に同意してくれた。続く数年間にこの資金援助でビードルのチームに加わった大学院生は、アディアン M.・サーブ、オーガスト H. (Gus)・ドールマン、フランクC.・ハンゲイト、タインT.・ベル、ベルナ・クーンラットとデビット・レグネリーの6人だった。
ビードルと同様ネブラスカの小さな農場で育ったサーブはリンカーンの農学科で学部生活を送り、ビードルの学部時代の助言者で友人でもあったカイム教授に誘われて研究の道に入った。ビードルに学べとカイムに薦められたサーブは、1941年にショウジョウバエのプロジェクトで研究できると期待してスタンフォードに大学院生として入学して来たが、 「実験室の大勢が、ネブラスカの農学科では見たこともない大量のオレンジ色のカビを扱っているのを見て」 驚いた48。スタンフォードはその点で特異で、実際に当時このカビを扱う実験室はまだ他には多くなかったであろう。ドールマンはイリノイ州出身で、ワバッシ・カレッジとイリノイ大学で生物学を学び、アカパンカビの仕事でチームが結成され始めた時にスタンフォードにやって来た。レグネリーは、ビードルの遺伝学の講義を受講した学部学生だったが、大学院生としてショウジョウバエの眼色の仕事を続ける熱意はなくカルテックへ移っていた。レグネリーもまたビードルのアカパンカビのセミナー講演に圧倒されてカビ・グループに加わるためにスタンフォードへやって来た一人だった。だが彼はすぐに軍隊に招集され、大学院の研究を完成するために帰って来ることができたのは1945年になってからだった。
学生達は研究課題を自ら生み出すべきだとビードルは強く信じていた。それは彼がコーネルとカルテックで育て上げられたやり方だった。ビードル自身が、学生時代にはもちろん研究の途上では資金の援助が約束されていたが、研究目標と研究計画を自分自身のデザインで決めるよう期待された一人だった。しかしスタンフォードでは、実際は主として学部学生とテクニシャンを配置した 「突然変異の探索作戦」 が実施され、それによって突然変異体の速やかな選抜が可能になり、全てが試験研究に供される状況だった。大学院生は、通常は先輩のポストドク研究員と共に仕事をしたが、いくつもの突然変異体を選抜してそれらの栄養要求性を決める仕事に従事した。彼らには、選抜した突然変異体の性質付け、欠損が単一遺伝子によるか否かの判定、染色体上へのマッピングに加えて、影響を受けた合成反応の同定が求められた。
ビードルとテータムは、それまで遺伝学者が始めることを回避して来た問題への新たな攻撃方法を開拓したのだった。遺伝子はまさに何をするのか?どのようにするのか?二人は最初の質問に対する答えを導くための実験的な手段を自分達が開発できたと信じた。彼らの実験は、生物のもつ代謝と発生の能力を作り上げる元となる無数の反応を触媒する酵素の生産を具体的に指令する点に遺伝子の役割があることを指摘していた。しかし、遺伝子と酵素がどのように関係しているのかはまだ不明であり、遺伝子が酵素の合成を 「案内する」 仕組みについては、これら初期の実験が示唆するものはまだなく、推測と議論の的として残っていた。いずれにしても、ビードルが短期間で結集したチーム・メンバーの優れた才能と献身的な努力、さらに必要な資源を提供したロックフェラー財団の賢明な判断のお蔭で、彼とテータムは遺伝子と酵素の関係について大発見を成し遂げることができた。こうして彼らは遺伝子と酵素の機能的な関係を究明する明確な方法を提供し得たが、それでもどのように遺伝子がそこで役割を果たすかが解明されるにはなお20年の歳月が必要だった。
1. E.L. Tatum. 1939. Development of eye colors in Drosophila: Bacterial synthesis of v+ hormone. Proc. Natl. Acad. Sci. 25: 486-490; E.L. Tatm and GWB. 1940. Crystalline Drosophila eye color hormone. Science 91: 458.
2. A. Butenandt, W. Weidel, and E. Becker. 1940. Kynurenine als Augenpigmentbildung Auslosendes Agens bei Insekten. Naturwissenschafen 28: 63-64.
3. E.L. Tatum and A. Haagen-Smit. 1940. Identification of Drosophila V hormone of bacterial origin. J. Biol. Chem. 140: 575-580.
4. Clarence P. BergからGWBへの手紙, January 14, 1947. Biological Division Records, Box 12.24, CIT.
5. H. Kikkawa, 1941. Mechanisms of pigment formation in Bombyx and Drosophila. Genetics 26: 587-607.
6. A. Butenandt, W. Weidel, and E. Becker. 1949. 3-oxy-kynurenine als cn-Gen Abhangiges Gleid im Intermediaren Tryptophan Stoffwechsel. Z. Naturforsch 4B: 242-244.
7. D. Ghosh and H.S. Forrest. 1967. Enzymatic studies on the hydroxylation of kynurenine in Drosophila melanogaster. Genetics 55: 423-431.
9. EphrussiからGWBへの手紙, October 23, 1939. Beadle Collection, Box 1.26, CIT.
10. 同上.
11. EphrussiからGWBへの手紙,February 19, 1940. Beadle Collection, Box 1.26, CIT.
12. ルイ・ラプキンはパリのパスツール研究所の有名な生物学者で、ユダヤ人研究者達をアメリカとその他の中立国へ脱出させ定住させる計画の立案に大きな役割を果たした.
13. Irene BarluetからP. Bergへの手紙, December 14, 1997およびロックフェラー財団記録RG 10F/S Recorder Cards Room 103 Red Drawers Draw #2.
14. GWB. Genetic control of the production and utilization of hormones. Proceedings of the 7th International Genetics Congress (suppl.). Cambridge University Press, 1941, pp. 58-62.
15. R.Scott-Moncrieff. The classical period in chemical genetics recollections of Muriel Wheldale Onslow, Robert and Gertrude Robinson and J.B.S. Haldane. Notes and Records of the Royal Society of London, 36, 1981, pp. 126-154.
16. A.E. Garrod. Inborn errors of metbolism. 2nd edition, Frowde and Hodder and Stoughton, 1923.
17. 1939年8月開催の第7回国際遺伝学会議での発表から論文の受理、編集と出版までに長い時間が経過しており、出版されたテキスト(1941年)がビードルが会議の際に実際に発表した内容と同一であるか不明.
18. GWB and E.L. Tatum. 1941. Experimental control of development and differentiation. Am. Nat. 75: 107-116.
19. GWB, Genetic control of production.
20. GWB. 1975. Biochemical genetics: Reflections. Three lectures, January 15-17, 1975掲載, The Edna H. Drane Visiting Lectureship, University of Southern California, School of Medicine. Norris Medical Library Archives W9B365t.
21. 講義ノートはJoshua Lederbergの提供,原文はNational Library of Medicine Archives所蔵.
22. A. Lwoff. 1938. Les Facteurs de Croissance Pour les Microorganismes. Ann. Inst. Pasteur 61: 580-617; B.C.J.G. Knight. 1936. Bacterial nutrition. His Majesty’s Stationary Office, pp. 1-182; R.J. Williams. 1941. Growth-promoting nutrilites for yeasts. Biol. Rev. 16: 49-80.
23. Sydney Raffel, インタビュー, August 31, 1999.
24. GWB. 1966. Biochemical genetics: Some recollections. Phage and the origin of molecular biology (ed. J. Cairns, G.S. Stent, and J.D. Watson), Cold Spring Harbor Laboratory Quantitative Biology. Cold Spring Harbor, New York, pp. 23-32掲載; ビードルはドッジによる遺伝交配の結果の誤った解釈を大学院生達が正さなければならなかったことをしばしば思い出して語った。ドッジは減数分裂で起こる組換えの時期に従って特定の交配で生じた子嚢胞子の遺伝的特徴がどのように変化するかを説明する新しい概念に馴染んでいないようだった。
25. ドッジが退職した時、ビードルは次のようなコメントを残している。 「ドッジ家とビードル家の先祖達は3世紀前に何かグロテクスな形で遭遇していたようだ」 。ドッジ家の先祖の一人は、魔女だと申し立てられた者が尋問を受けるために集められたと記録されているサレムで、魔女裁判による焼身刑で亡くなっているが、ビードルの8世代前の従兄弟はサレムでビードル酒場を経営していたようである。ビードルは、もしかすると 「ドッジ家の親類もビードル酒場へ連行されたのでは」 と疑ったようだ。Beadle Collection, Box 3.9, CIT.
26. Garland E. AllenからJoshua Lederbergへの手紙, September 6, 1979. Joshua Lederberg提供;GWB. 1959. Genes and chemical reactions in Neurospora. Science 129: 1715-1719; C.C. Lindegren. 1942. The use of fungi in modern genetical analysis. Iowa State Coll. J. Sci. 16: 271-290; C.C. Lindegren and G. Lindegren. 1941. X-ray and ultra-violet mutations induced in Neurospora. J. Hered. 32: 405-412.
27. A.H. Sturtevant and GWB. An introduction to genetics. W.B. Saunders, Philadelphia and London, 1940.
28. GWBからBernard O. Dodgeへの手紙, February 27, 1941. New York Botanical Garden Library. Dodge Collection.
29. 同上.GWBからDodgeへの手紙, March 13, 1941.
30. F. Kogl and N. Fries. 1937. Uber den Einfluss von Biotin, Aneurin und Meso-Inositol Wachstrum Verschiedener Pilzarten. Hoppe-Seyl. Z. 249: 93-106.
31. 植物園のドッジの同僚W.J. Robbinsがビオチンは精製綿から熱水抽出で容易に得られる事実を発見して以来ビオチン不足は解消されていた.
32. H.J. Muller. 1927. Quantitative methods in genetic research. Am. Nat. 61: 407-419.
33. David Regnery,インタビュー,July 3, 1996.
34. GWB and E.L. Tatum. 1945. Neurospora II. Methods of producing and detecting mutations concerned with nutritional requirements. Am J. Bot. 32: 678-686.
35. GWBからCarl C. Lindegrenへの手紙,July 25, 1941. Beadle Collection, Box 5.10, CIT.
36. GWBからDodgeへの手紙、August 11, 1941. New York Botanical Garden, Dodge Collection.
37. GWB and E.L. Tatum. 1941. Genetic control of biochemical reactions in Neurospora. Proc. Natl. Acad. Sci. 27: 499-506.
38. N.H.Horowitz. George Wells Beadle: Biographical memoirs of the American Philosophical Society Year Book, American Philosophical Society, 1995.
39. Sydney Raffel, インタビュー,August 31, 1999.
40. B. EphrussiからGWBへの手紙,August 22, 1941. Beadle papers, Box 1.26, CIT.
41. GWBからEphrussiへの手紙,November 17, 1941. Beadle papers, Box 1.26, CIT.
42. GWBからWarren Weaverへの手紙,November 28, 1941. 205D, RG 1.1, Box 10, Folder 141, RFA.
43. 同上.
44. Frank B. Hansonがビードルと面会した折の日記, Dec 15, 17, and 18, 1941. 205D, Box 10, Folder 141, RFA collection.
45. Hansonの日記.September 4 and 5, 1937. 205D, RG1.1, Box 7, Folder 88, RFA.
46. HansonからGWBへの電報,1942. 205D, RG1.1, Box 10, Folder 141, RFA.
47. GWB. 1978. Recollections. Annu Rev, Biochem. 43: 1-13.
48. A.M. Srb. 1973. Beadle and Neurospora. Some recollections. Neurospora Newslett. pp. 8-9.
| <<前の章 |
TOP |
次の章>> |
|
Copyright (C)
2014 Chiharu Nakamura All Rights Reserved.
|
|