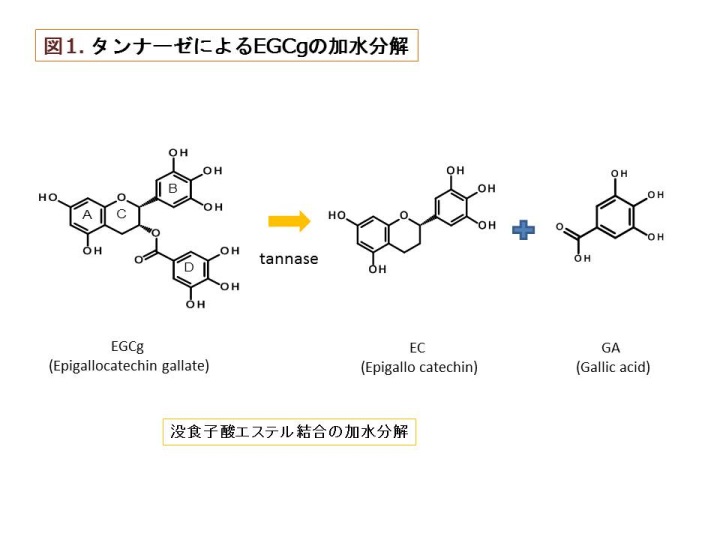
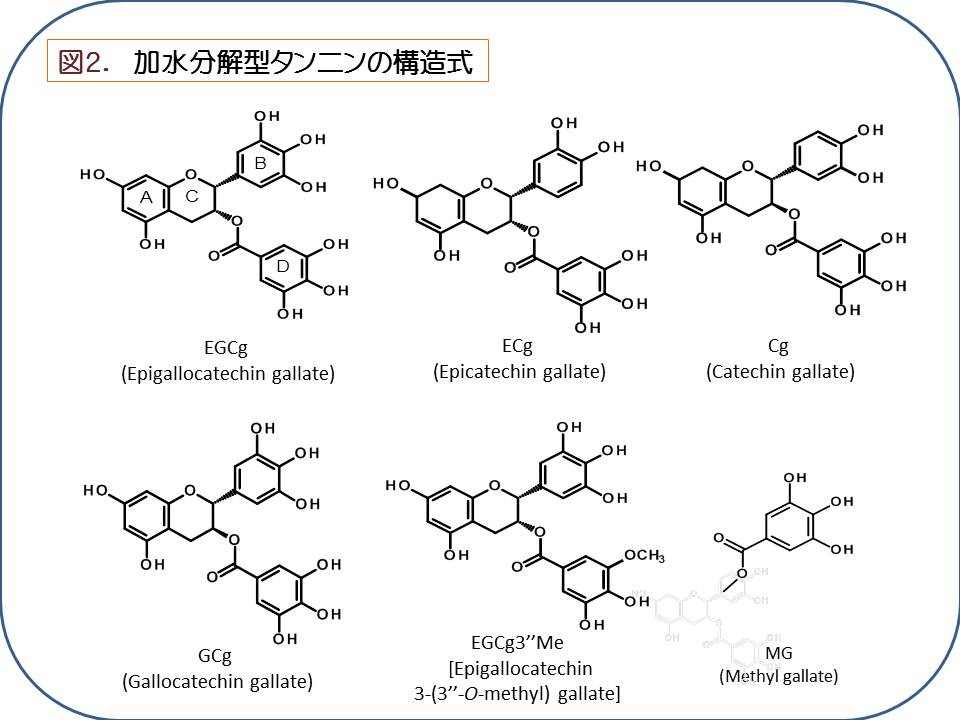
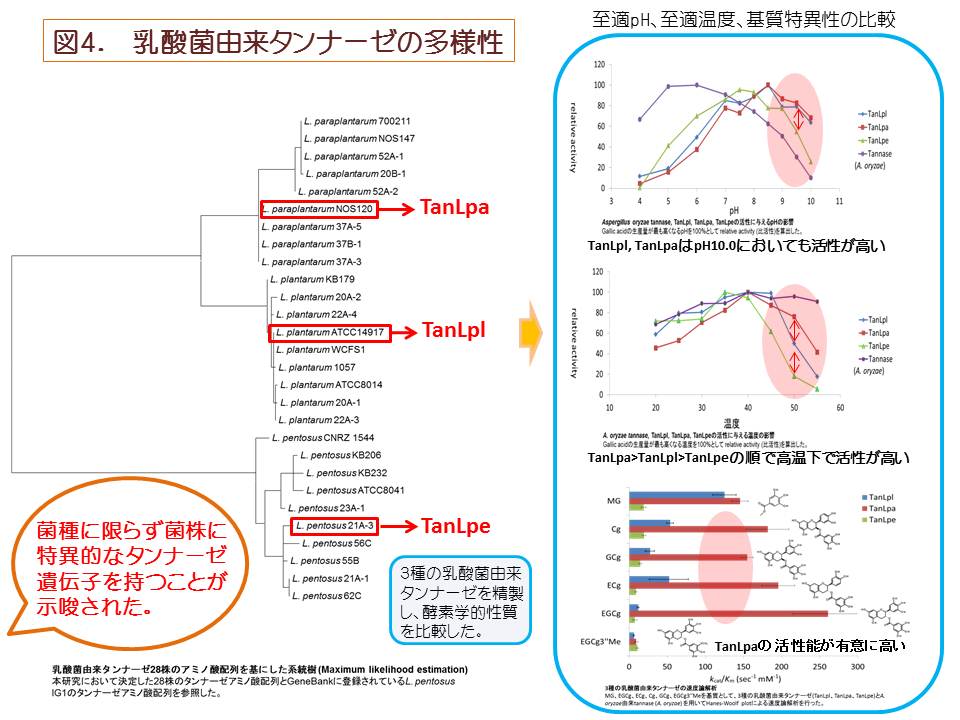
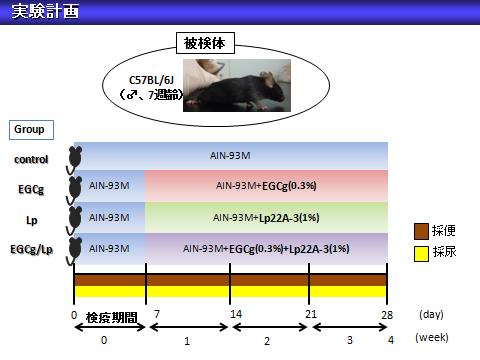
(1)タンナーゼ産生性乳酸菌に関する研究
タンナーゼ(tannin acyl hydrolase, EC 3.1.1.20)は、加水分解型タンニンの構造的な核である多価アルコールとフェノール酸類 との間に形成されているエステル結合を加水分解する酵素である。加水分解型タンニンは植物界に普遍的に存在し、抗菌活性を有する ポリフェノール化合物であり、タンパク質や重金属等と結合して通常の消化酵素では消化できない難容性の塩を形成する性質を有する。 ある種のカビや細菌はタンナーゼを産生し、これによって上記の難容性塩を消化して植物体から栄養やミネラルを奪取していると考えられている[1]。 過去に、本研究室の黒髭先生(大澤 朗教授)はコアラが盲腸にタンナーゼを産生する細菌(Lonepinella koalarum)を戦略的に宿し、主食であるユー カリの葉のタンニン成分とタンパク質の複合体を本菌が分解することによって、窒素源の吸収に重要な役割を担っていることを報告した[2, 3]。 タンナーゼが分解する加水分解型タンニンの一種として例えば、緑茶に含有するカテキン類がその一種である。これまでに、真菌類由来タンナーゼ についてはその酵素学的性質が解析され、産業的にも緑茶製造過程において生じるクリームダウン(白濁沈殿)を分解するのに広く利用されている。 タンナーゼの加水分解の例として、加水分解型タンニンの一種である没食子酸エピガロカテキン(epigallocatechin gallate、EGCg)がタンナーゼに よって加水分解される過程を図1に示した。EGCgは、タンナーゼによる加水分解によりエピガロカテキンガレート(epigallocatechin, EGC)と没食子酸 (Gallic acid, GA)に遊離する。また、メチルガレート(Methyl gallate, MG)、プロピルガレート(propyl gallate, PG)、カテキン類の没食子酸 エピカテキン (epicatechin gallate、ECg)などの没食子酸エステル結合を加水分解し、没食子酸を遊離させる(図2)。 これまでに本研究室では、乳酸菌の一種Lactobacillus plantarumにおいてタンナーゼの産生を確認し、タンナーゼ遺伝子の同定とその性質の解析が なされてきた[4, 5, 6]。そこで本研究ではLactobacillus plantarumと漬物由来で近縁な L. paraplantarum、L. pentosus )の乳酸菌由来タンナーゼ の多様性と酵素学的性質について研究を行った。 本研究では、まず 同3種の乳酸菌由来タンナーゼのアミノ酸配列を同定し、さまざまな由来からなる28菌株で比較した。するとTanLplとTanLpaの アミノ酸配列の相同性は88%であり、TanLpeとTanLpl、TanLpaとの相同性はそれぞれ72%、73%であった。これまでに乳酸菌由来タンナーゼは構 造解析がなされており、酵素活性に重要なアミノ酸であるAsp421-Lys343-Glu357は3種の乳酸菌由来タンナーゼでいずれも保存されていた(図3)[7]。 3種の乳酸菌(L. plantarum、L. paraplantarum、L. pentosus)は16S rRNA オペロンの塩基配列や生理生化学的性状が似通っており、非常に近縁な種である。 このような近縁な種においてもアミノ酸配列、乳酸菌由来タンナーゼが変化を起こしやすく、自然界において多様な構造が存在することが推察される。 乳酸菌由来タンナーゼに多様な構造が存在する理由として、自然界で植物は自身の体内に多様な構造の抗菌物質を産生し 、防御機構を形成している と考えられており、酵素学的性質を多様化させることで乳酸菌が植物の防御機構に対抗していることが推測される。 次に、至適温度、至適pHの乳酸菌由来タンナーゼについて乳酸菌が実際にどのような環境下で利用しているか、本酵素を利用できる環境を推測した。 3種の乳酸菌由来タンナーは共通してpH 7.0-pH 9.0において安定であり、至適温度は40℃付近であることが示唆された。自然界において乳酸菌が生息し、 且つこのような条件の環境として、糞便や食料残渣の堆肥が考えられる。一般に堆肥化の過程において糖類やアミノ酸などの有機物が分解され、 アンモニアが発生し、アルカリ性の環境になり(pH 7.0-pH 9.0)、同時に微生物の発酵熱で40℃付近まで温度が上昇することが知られている[8]。 このような堆肥のアルカリ性の環境下ではBacillus属やClostridium属の生育が優勢であり[9]、至適な生育pHが弱酸性(pH 5.0-pH 6.0 )である乳酸菌は有意に生息できる環境ではないと考えられるが、一方で、L. plantarumやL. pentosusはアルカリ性(pH 8.5-pH 8.9まで) の環境においても、生存できるような耐性をもつことが報告されており[10]、このような環境下に適応して生育していることが推察される。 続いて、さまざまな構造の没食子酸型カテキン類を用いて速度論解析を行い、3種の乳酸菌由来タンナーゼの基質特異性の差異を検証した。 3種の乳酸菌由来タンナーゼ全てにおいて、特に没食子酸カテキン類を基質とした場合、Km値はそれぞれ同程度の値を示すが、触媒効率を示すkcat値が TanLpaはTanLpl、TanLpeと比較して 5倍から10倍程度高く、TanLpaは3種の乳酸菌由来タンナーゼの中でも没食子酸型カテキン類に対する分解能に 優れていることが示唆された。このような触媒効率が高い酵素を持つことは、環境中に存在する限られた基質を他の種よりも速く分解し、 有意に栄養を摂取できると推測されるため、生存に有利であると考えられる。EGCg3''Meを基質とした反応ではEGCgとの反応と比較して、 3種の乳酸菌由来タンナーゼに共通して親和性に変化はなかったが、触媒活性が低下し、有意に活性能が低下した。これはEGCg3''Meの構造がEGCgと異なり 、没食子酸の3位のヒドロキシ基がメトキシ基であることで、乳酸菌由来タンナーゼによるエステル結合の加水分解反応が妨げられているものと考えられる。 一方、真菌類由来タンナーゼの場合は活性能が落ちないことから乳酸菌由来タンナーゼの方が真菌類由来タンナーゼよりも基質特異性が高いことが示唆された。