 |
 |
|
|
|
|
(a) We performed resonance Raman spectroscopic analyses on the reduced-CO forms of hemoglobin and myoglobin and identified Fe2+-CO stretching vibration (507 cm-1) and Fe2+-C-O bending vibration (578 cm-1) for the first time[7]. We also conducted resonance Raman studies on azide (N3-) and cyanide (CN-) bound forms of hemoglobin and myoglobin in oxidized state and identified Fe3+-N3 and Fe3+-CNstretching vibrations for the first time[4, 5, 8]. Further, in colaboration with Dr. Kitagawa, we first identified the Fe2+-Ne(His) stretching vibration [1]. These studies made great influences in the field of "Resonance Raman analyses of heme-ligand interaction" in the hemoprotein studies.
(b) Cobalt-substituted hemoglobin and myoglobin are well-known as good models for the iron-containing hemoglobin and myoglobin for the study of dioxygen-binding at the heme center. We studied dioxygen-binding to cobalt-substituted hemoglobin and myoglobin using resonance Raman spectroscopy and identified bound O-O stretching and Co-O2 stretching vibrations for the first time. These results explained the unusual infrared O-O stretching band and the nature of dioxygen binding in iron-containing hemoglobin and myoglobin [6, 9].
(c) Azide ion can bind manganese-substituted myoglobin. We found that the coordinated azide ion could be photolyzed leading to a high-valent Mn(V)-nitrido species characterized by a Mn-N stretching resonance Raman band at 1010 cm-1 and its first overtone mode at 2006 cm-1[17, 26].
(2) Enzymatic and Physicochemical Analyses of the Steroid Hormone Hydroxylase Systems in Adrenal Cortex
(a) Adrenal cortex mitochondria contain cytochrome P450scc responsible for the cholesterol side-chain cleavage reaction, the rate-limiting step of steroid hormone biosynthesis. We identified an Fe2+-CO stretching vibration (477 cm-1) of its CO-reduced form by the resonance Raman spectroscopy for the first time. Simultaneously, a bound C-O stretching vibration was identified at 1953 cm-1. We previously found a clear inverse relationship between the Fe2+-CO stretching vibration and the bound C-O stretching vibration among a hemoprotein family containing a histidine residue as the heme 5-th axial ligand. Based on this, we proposed that cytochrome P450 family constituted a distinct group due to the coordination of a cysteinyl thiolate ligand to the heme iron [11, 13, 15].
(b) Mitochondrial-type cytochromes P450 (i.e., cytochrome P450scc and cytochrome P45011b, etc.) receive electron equivalents from a [2Fe-2S] iron-sulfur protein, adrenodoxin. The electron equivalents are absolutely required for the dioxygen activation and the hydroxylation reactions. We identified the adrenodoxin-binding site on the cytochrome P450scc sequence for the first time by utilizing a specific chemical modification with PLP. The sequence (Lys377-Lys381) is well-conserved among the mitochondrial-type cytochrome P450 subfamily [21] and, later, was confirmed as an adrenodoxin-binding sequence by a site-directed mutagenesis study.
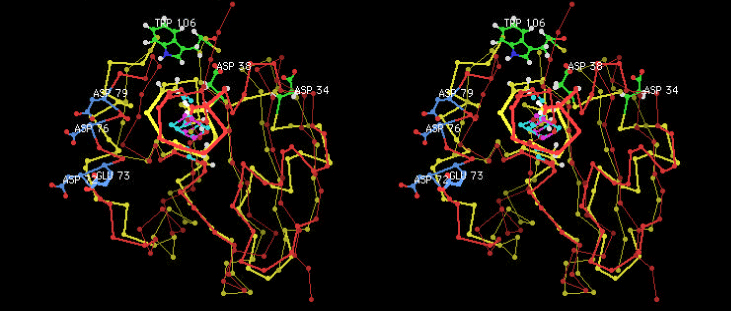
(c) The specific interaction between adrenodoxin and cytochrome P450scc is believed to be electrostatic. We analyzed the nature of the interaction by employing EPR spectroscopy. The results showed a distinct difference in the nature of the interaction in comparison with the system of cytochrome P450cam and putidaredoxin. This difference might be originated from the interacting amino acid residues of two ferredoxins as shown below (Fig. 1) [64].
(d) EPR spectroscopic analysis of cytochrome P450scc(Fe2+)-NO complex showed that substrates and its reaction intermediates in the cholesterol side-chain cleavage reaction caused dramatic changes on the EPR spectra. These spectral changes were considered as due to consecutive conformational changes at the active site to proceed a series of the side-chain cleavage reactions properly [16, 19].
(e) We found that binding of reduced adrenodoxin to cytochrome P450scc(Fe2+)-phenylisocyanide complex caused a dramatic spectral change to occurr in the heme absorption spectrum (Soret band region). This spectral change was interpreted as the tertiary structural change around the heme moiety to make an electron transfer reaction from iron-sulfur center to heme active site [22].
(f) We purified cytochrome P45011b from bovine adrenal and analyzed by utilizing various biophysical and enzymological techniques. This cytochrome P450 exists in inner membrane of adrenal cortex mitochondira and is responsible for the biosynthesis of mineral corticoids and sugar corticoids [24].
(g) We purified cytochrome P450c21 and analysed by utilizing various biophysical and enzymological techniques. This cytochrome P450 exists in microsomal fraction of adrenal cortex and is responsible for the biosynthesis of mineral corticoids and sugar corticoids. We found further that this enzyme has a previously unkonwn steroid 20b-oxidase activity [32, 43, 72].
 |
||||
| Fig. 1 Differences in interactions between the P450scc-adrenodoxin and the P450cam-putidaredoxin systems ( stereo view .) (Sticks and balls in yellow and yellowish green indicate P450cam, whereas those in red and blue indicate P450scc.) |
||||
(3) Structural and Functional Analyses of the Respiratory Heme-Copper Terminal Oxidases Using Molecular Biological Techniques
(a) To clarify the structure of heme-copper binuclear mitochondrial-type cytochrome c oxidase, we analyzed the CO-, azide-, and cyanide-bound forms of cytochrome c oxidase using FT-IR and EPR spectroscopies. We proposed a structural model based on thses measurements [31, 33, 34]. Especially, we found clear evidence that azide and cyanide bind to the heme-copper center as a bridging ligand between heme and copper and further the mode of binding varies dependeing on the redox levels of the metal centers.
(b) We purified bo- and ba-type ubiquinol oxidase from E. coli and A. aceti, respectively. These enzymes belong to heme-copper terminal oxidase family and expected to have similar mechanism to pump protons across the biomembranes with that of mitochondrial cytochrome c oxidase. We did extensive investigation on these enzyme using FT-IR, resonance Raman, and EPR spectroscopies [35, 45, 49, 51, 57]. We found that these ubiquinol oxidase have very similar active site structures with those of cytochrome c oxidase and, therefore, can be good models for the investigation of the mechanism of redox-linked proton pumping [35].
(c) Heme-copper oxidases have the most important role for the biological energy transduction by their proton pumping activities. We investigated the active site structure of various site-directed mutants of E. coli bo-type ubiquinol oxidase using FT-IR, resonance Raman, and EPR spectroscopies. We proposed a plausible structural model at the heme-copper binuclear active site based on these studies [38, 41, 48, 52, 54].
(4) Structural and Functional Analyses of the Heme-Heme Terminal Oxidase
(a) E. coli has a respiratory terminal oxidase, named bd-type ubiquinol oxidase, different from bo-type ubiquinol oxidase. This enzyme has a completely different amino acid sequence and different physical properties from those of the heme-copper oxidase. We investigated the active site structure of this bd-type ubiquinol oxidase using various biophysical techniques. Our FT-IR study on the CO-bound air-oxidized form showed a very different characteristics (a high C-O stretching frequency and a low Fe2+-CO stretching frequency). We clarified further that the air-oxidized form is actually a dioxygen-bound form based on the replacement with nitric oxide[37].
(b) We found that the purified form of bd-type ubiquinol oxidase has an unique active site consisting of two hemes b based on a heme content analysis, visible absorption spectroscopy, and FT-IR analyses on cyanide- and azide-bound forms for the first time [44, 58, 59].
(c) We identified that the heme d axial 5-th ligand is a nitrogen atom based on the EPR spectroscopic analysis of the NO-bound form. Heme d center is considered as the dioxygen-binding site at the heme-heme binuclear center [46].
(5) Analyses of the Electron Transfer System in Neuroendocrine Vesicles (Cytochrome b561 and PAM)
(a) Neuroendocrine vesicles contain various neurotransmitters including noradrenaline, adrenaline, amidated neuropeptides. In the vesicular membranes, cytochrome b561 exists as a transmembrane protein and catalyzes transmembrane electron transfer from cytosolic ascorbate (AsA) to intravesicular monodehydroascorbate (MDA) radical. This electron transfer reaction is absolutely required for the intravesicular copper-containing monooxygenases, such as dopamine b-hydroxylase (DBH) and peptidylglycine a-hydroxylating monooxygenase (PAM), and is essential for the biosynthesis of neurotransmitters. We succeeded to purify cytochrome b561 from chromaffin vesicles of bovine adrenal medulla and found that the purified cytochrome contained two heme b per molecule by a heme content analysis and EPR spectroscopy [50].
(b) We cloned cytochrome b561 cDNA from various animals. Based on the nalysis of deduced amino acid sequences, we proposed that two well-conserved sequences, possibly an ascorbate (AsA)- and an monodehydroascorbate (MDA)-binding sites reside on the cytosolic and intravesicular sides, respectively. Additionally, there were 5 fully conserved His residues, possible heme ligands on both sides of the membranes [55] .
(c) We revealed that the two heme centers have distinct physiological roles, electron acceptance from ascorbate (AsA) and electron donation to monodehydroascorbate (MDA) radical, by a pulse radiolysis technique [56].
(d) We found that DEPC-treatment of cytochrome b561 in the oxidized state caused a significant inhibition of electron acceptance from ascorbate. We identified the DEPC-modification sites by an MALDI-TOF-MS technique[60]. We further found that the inhibition in the electron acceptance by DEPC could be protected by the inclusion of ascorbate during the DEPC-treatment, suggesting the presence of physiological ascorbate-binding site on the molecule [62].
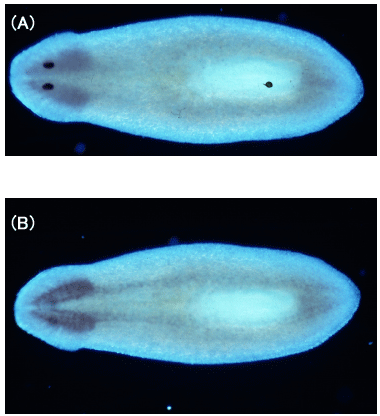
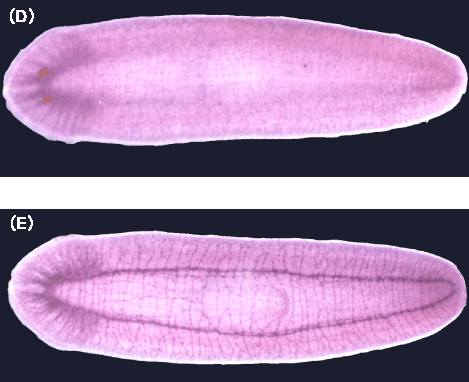
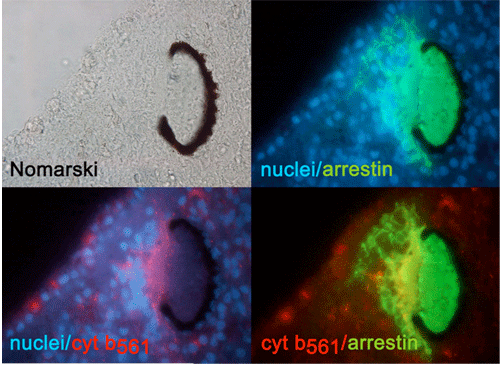
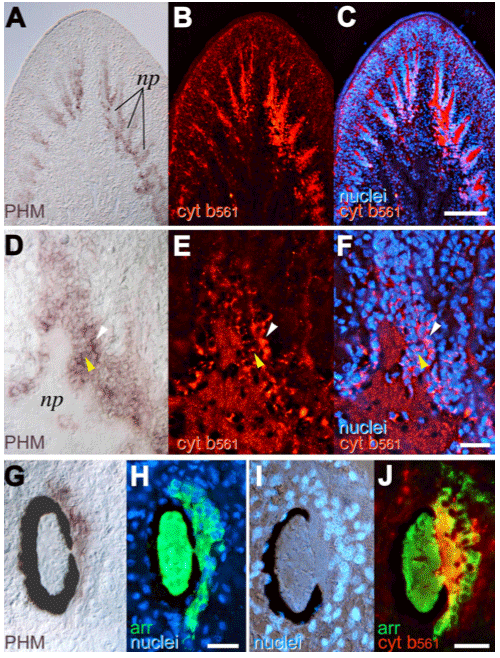
(e) We analyzed cytochrome b561 and PAM expressions in planarian body with cDNA cloning, in situhybridization, and immunohistochemical staining and found that these proteins were expressed along the planarian nervous system [65].
(f) We made extensive MALDI-TOF-MS analyses on the purified bovine cytochrome b561 . We identified and isolated N-terminal tryptic peptide with a reversed-phase HPLC and AARE-treatment. We found that this peptide has an acetyl group at the intiation Met residue based on the Edman amino acid sequence analysis and the MALDI-TOF-MS analysis. There was no other post-translational modification to ocurr, like the acylation with a fatty acid group, as previously suggested [66].
(g) We conducted a stopped-flow study on the reaction of AsA with purified cytochrome b561 in the detergent-solubilized state for the first time. The time-course of the reduction of oxidized cytochrome b561 with AsA could not be fitted with a single exponential but with a linear combination of at least four exponential functions. The fastest phase, which was assigned to the first 1-electron donation from AsA to heme b on the extravesicular side, was further analyzed by transient phase kinetics employing a two-step Bi-Uni sequential ordered mechanism. The result showed Ks=2.2 mM for AsA at pH6.0. At a region below pH5.5, there was a significant lag before the reduction of hemes b occurred. This time-lag was interpreted as due to a pH-dependent transient state before the first electron transfer to take place. The fastest phase was completely lost by N-carbethoxylation of heme-coordinating histidyl residues (His88 and His161) and Lys85 upon treatment with DEPC. Presence of AsA during the treatment inhibited the N-carbethoxylation of the histidyl residues and, thereby, restored the final reduction level of hemes b. But the reduction rate was still only one-twentieth of the native form. This result suggested an important role of the conserved Lys85 for the interaction with AsA [67].
(h) Purified cytochrome b561 was reconstituted into AsA-loaded vesicles. The intravesicular AsA was able to reduce external ferricytochrome c. The reduction of cytochrome c was dependent on the presence of cytochrome b561 in the vesicle membranes. The topology of the reconstituted cytochrome b561 in the vesicle membranes was examined by treatment with trypsin followed by SDS-PAGE and MALDI-TOF-MS analyses. We found that cytochrome b561 was reconstituted into the membranes in an inside-out orientation irrespective of the modification with DEPC. The addition of a soluble form of DBH to the external medium resulted in the successful reconstitution of the hydroxylation activity towards tyramine, an analogue of dopamine. This activity was enhanced upon the addition of ferricyanide as a mediator between cytochrome b561 and DBH [68].
(i) Treatment of oxidized cytochrome b561 with DEPC caused a downshift of midpoint potential for the lower component and this shift was protected by the presence of AsA during the treatment. Present EPR analyses showed that, upon the treatment, the gz=3.69 heme species was converted to a non-reducible form with AsA, although its gz-value did not show any appreciable change. The treatment did not exert any influence on the other heme (the gz=3.13 species) at all. Raman data indicated that two heme b centers adopt six-coordinated low-spin state, both in the reduced and oxidized forms. There was no significant effect of the DEPC-treatment on the Raman spectra in both forms but the reducibility with AsA differed significantly between the two hemes upon the treatment. Addition of ferrocyanide enhanced both reduction rate and final reduction level of the DEPC-treated cytochrome b561 when AsA was used as a reductant. This observation suggested that ferrocyanide scavenges MDA radical produced by a univalent oxidation of AsA and, thereby, increases both the reduction rate and the final reduction level of the heme center on the intravesicular side of the DEPC-treated cytochrome. This result further clarified the physiological role of this heme center as the electron donation to MDA radical [69].
 |
|||||||||||||||
| Fig. 2 Transmembrane structural model of bovine cytochrome b561 There are six transmembrane α-helices. Those are connected with short hydrophilic loops. Both N- and C-terminus of the protein are considered to be on the extravesicular side. Two well-conserved sequences in animal species, 5 totally-conserved and one well-conserved His and one conserved Met residues are indicated. Further, well conserved basic amino acid residues are indicated by +. These might have important roles for the transmembrane electron transfer from extravesicular ascorbate to intravesicular monodehydroascorbate radical. |
|||||||||||||||
 |
|||||||||||||||
| Fig. 3 Distribution of cytochrome b561-mRNA along the planarian nervous system (in situ hybridizaion) (A)Dorsal view, (B)ventral view. |
|||||||||||||||
 |
|||||||||||||||
 |
|||||||||||||||
| Fig. 4 Ditribution of cytochrome b561 protein along the planarian nervous system immunostaining with anti-planarian cytochrome-b561 antibodies. (C) Western blotting;the 27.5KDa band indicates the planarian cytochrome b561, (D) Dorsal view, (E) ventral view. |
|||||||||||||||
 |
|||||||||||||||
| Fig. 5 Planrian eye Cytochrome b561 and arrestin are stained with respective specific antibodies arrestin : protein marker of eye ( nuculei are stained with Hoechst 33342. ) |
|||||||||||||||
 |
|||||||||||||||
| Fig. 6 Planarian brain and eyes Planarian brain and eye, stained with PHM (in situ hybridization) and cytochrome b561 (antibody) PHM : corrseponding to the monooxygenase domain of PAM |
|||||||||||||||