|
 |
|
|
|
|
|
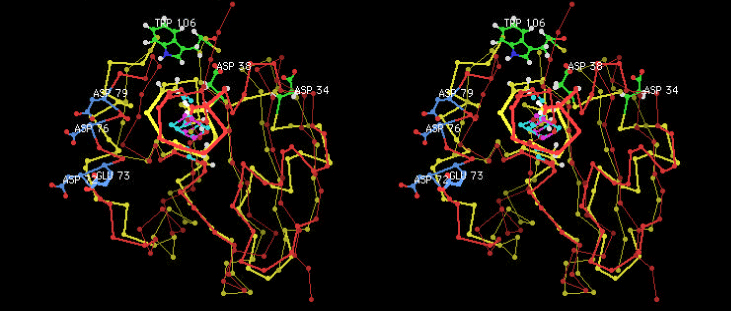
(1) ヘモグロビン・ミオグロビンの共鳴ラマン散乱による解析 (a) ヘモグロビン・ミオグロビンの還元型ヘム鉄に一酸化炭素(CO)が結合した状態を共鳴ラマン散乱法により解析し、Fe2+-CO 伸縮振動 (507 cm-1) 、Fe2+-C-O 変角振動 (578 cm-1)を初めて同定した[7]。またアザイド(N3-)、シアン(CN-) 等と酸化型ヘム鉄との結合を共鳴ラマン散乱法により解析し、Fe3+-N3 伸縮振動、Fe3+-CN伸縮振動等を初めて同定した[4, 5, 8]。また、還元型ミオグロビンにおけるFe2+-Ne(His)の伸縮振動を初めて同定した[1]。これらの仕事により「ヘム鉄への配位子結合の共鳴ラマン散乱による解析」というヘムタンパク質研究の新しい分野を切り開いた。 (b) 鉄をコバルトに置換したヘモグロビン・ミオグロビンにおいて、コバルトヘムに結合した酸素分子のO-O 伸縮振動バンドを共鳴ラマン散乱法により同定し、赤外分光スペクトル中で見られるこのバンドの異常な振る舞いの原因と、天然ヘモグロビン・ミオグロビンでの酸素結合様式を推定した [6, 9]。 (c) マンガン置換ミオグロビンにおいてマンガンヘムに結合したアザイドが光分解され、高酸化状態Mn(V)=N(ニトリド)が形成されること、そしてその伸縮振動(1010 cm-1)及びその倍音(2006 cm-1)が共鳴ラマン散乱法によりとらえられることを明らかにした[17, 26]。 (2) 副腎皮質ステロイドホルモン水酸化酵素系の酵素化学的・物理化学的解析 (a) 副腎皮質ミトコンドリアに存在するチトクロムP450scc(コレステロール側鎖切断酵素)のFe2+-CO 伸縮振動(477 cm-1) と結合C-O伸縮振動(1953 cm-1) を共鳴ラマン散乱法によって初めてとらえた。その振動数と結合C-O伸縮振動数の解析より、チトクロムP450がこれまで知られているヒスチジン・イミダゾール基を第5配位子とする通常のヘムタンパク質とは異なる新しいグループ(システインを第5配位子とする)を形成していることを明らかにした [11, 13, 15]。 (b) ミトコンドリア型チトクロムP450は基質の水酸化反応に必要な電子等量を鉄硫黄タンパク質・アドレノドキシンより受け取っている。PLP による特異的化学修飾法により、チトクロムP450scc 上のアドレノドキシン結合部位(Lys377-Lys381)を初めて明らかにした[21, 69]。この部位はミトコンドリア型チトクロムP450において非常に良く保存されており、後に部位特異的変異体の解析によりアドレノドキシン結合部位として確認されている。チトクロムP450sccとアドレノドキシンとの相互作用をP450camとプチダレドキシンとの相互作用と比較解析し、2つのシステムでの違いを明らかにした[64]。 (c) 還元型チトクロムP450scc と一酸化窒素(NO)との複合体のEPRスペクトルの解析より、コレステロール側鎖切断酵素反応の中間体を始めとする各種基質の存在下で、そのスペクトルが大幅に変化することを明らかにした。これは連続した水酸化反応により側鎖切断反応を行うための酵素活性中心における構造変化に対応したものと考えられる [16, 19]。 (d) 還元型チトクロムP450scc-フェニルイソシアナイド複合体に還元型アドレノドキシンが結合すると、ヘムの可視吸収スペクトル変化が起こることを初めて見出した。この変化はアドレノドキシンがいったん還元型チトクロムP450sccに結合した後、鉄硫黄クラスターよりチトクロムP450sccヘム鉄へ電子伝達をするために必要な3次構造変化に対応したものと思われる[22]。 (e)副腎皮質ミトコンドリアに存在し、鉱質コルチコイド・糖質コルチコイドの合成に関与するチトクロムP45011βの生物物理学的・酵素化学的解析を行った[24]。 (f)副腎皮質ミクロソームに存在し、鉱質コルチコイド・糖質コルチコイドの合成に関与するチトクロムP450c21の生物物理学的・酵素化学的解析を行った結果、この酵素がこれまで知られていなかったステロイド20β-オキシダーゼ活性を持つことを明らかにした[32, 43, 72]。
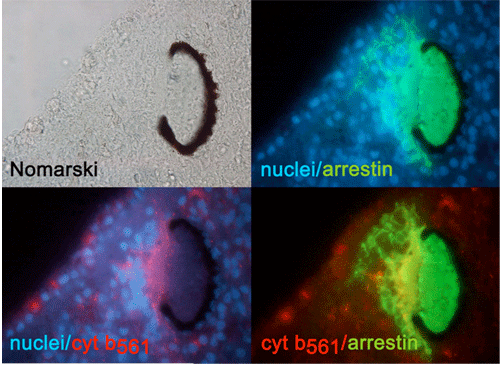
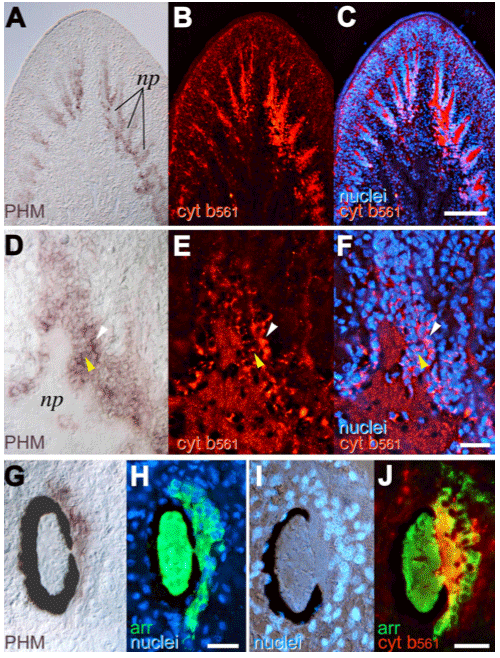
(3) 分子生物学的手法による呼吸鎖ヘム-銅型末端酸化酵素の構造と機能の解析 (a) ミトコンドリア型チトクロムc酸化酵素 のヘムー銅複核中心における構造を明らかにするために、一酸化炭素、アザイド、シアンをプローブとして赤外分光法、EPR法により精密な解析を行い、複核中心へのこれら呼吸阻害剤の結合様式を明らかにした[31, 33, 34]。特に、アザイド、シアンが複核中心において架橋型配位子として存在していることを明らかにし、金属中心の部分還元に伴い、その結合様式が変化することを明らかにした。 (b) ミトコンドリア型チトクロムc酸化酵素と同じヘムー銅末端酸化酵素ファミリーに属し、大腸菌・酢酸菌細胞膜に存在するbo, ba型ユビキノール酸化酵素を精製し、赤外分光法、EPR法等による複核中心の構造解析を行った[35, 45, 49, 51, 57]。ユビキノール酸化酵素はヘムー銅末端酸化酵素を研究する上で非常に良いモデルとなることを示した[35]。 (c) 生体膜中に存在し生体エネルギー変換において重要な位置を占めるヘムー銅末端酸化酵素の持つプロトンポンプ機構を明らかにするため、大腸菌 bo型ユビキノール酸化酵素の部位特異的変異体を作製し、赤外分光、共鳴ラマン散乱、EPR法による解析を行い、ヘムー銅末端酸化酵素活性中心付近の構造モデルを提出した[38, 41, 48, 52, 54]。 (4) ヘム-ヘム型末端酸化酵素の構造と機能の解析 (a) ヘムー銅末端酸化酵素とは機能・構造上異なるが、呼吸鎖末端酸化酵素であるシトクロムbd型ユビキノール酸化酵素を大腸菌細胞膜から精製し、生物物理的手法により解析した。一酸化炭素結合型酵素が通常のヘムタンパク質とは非常に異なった性質(高いC-O伸縮振動数、低いFe2+-CO伸縮振動数)を持っていることを明らかにし、精製酵素が実際には酸素結合型酵素であることを一酸化窒素との置換により明らかにした[37]。 (b)精製酵素のヘム含量、分光解析とシアン、アザイド結合型酵素の赤外分光測定により、活性中心が2つのヘムよりなるユニークな複核中心を形成していることを初めて明らかにした[44, 58, 59]。 (c)一酸化窒素(NO)結合型酵素のEPRスペクトル解析により、複核活性中心において、酸素分子が結合するヘムdの第5配位子は窒素原子であることを初めて明らかにした[46]。 (5) 神経内分泌小胞膜におけるシトクロムb561, PAMを中心とする電子伝達系の解析 (a) 神経伝達物質(ノルアドレナリン・アドレナリンなどの生体アミン、アミド化神経ペプチド)の貯蔵と放出に携わっている神経内分泌小胞には膜貫通型シトクロムb561が存在し、細胞質アスコルビン酸から小胞内モノオキシゲナーゼ(ドーパミン-β水酸化酵素・ペプチジルグリシンアミド化酵素(PAM))への電子伝達を行っている。この電子伝達はこれら神経伝達物質の合成に必須である。シトクロムb561を高純度に精製する方法を確立し、1分子中に2個のヘムBが結合していることを見出した[50]。 (b) 各種動物のシトクロムb561 cDNA クローニングよりアミノ酸配列を明らかにし、アスコルビン酸・セミデヒドロアスコルビン酸結合部位と思われる2つの保存領域が細胞質側、小胞膜内側にそれぞれ存在すること、また2つのヘムの結合部位も保存領域近くに存在する事を提唱した[55]。 (c) それぞれのヘムBがアスコルビン酸・セミデヒドロアスコルビン酸との電子伝達反応において、電子受容、電子供与という特異的役割を持つことをパルスラジオリシス法により解明した[56]。 (d) DEPCによる特異的化学修飾によりそれぞれの反応を特異的に阻害できることを明らかにし、その阻害部位をMALDI-TOF-MS法により特定した[60]。さらにDEPCによる阻害がアスコルビン酸の存在で阻止できることからチトクロム b561上に生理的アスコルビン酸結合部位が存在することを明らかにした[62]。 (e) 扁形動物プラナリアでのシトクロムb561とPAMの神経特異的発現をcDNAクローニング、in situ hybridization、組織免疫染色等により解析した[65]。 (f) 精製したウシシトクロムb561のNH2末端付近のペプチドを詳細に解析した結果、末端のMet残基がアセチル化されていることを明らかにした。しかし、以前に提唱されていた脂肪酸による翻訳後修飾は存在しないことがわかった[66]。 (g) 精製・可溶化した酸化型シトクロムb561へのAsAからの電子伝達反応をstopped-flow法により解析した。ヘムの還元反応曲線は最小でも4つの指数関数の和でなければ近似できなかった。これは電子がAsAから細胞質側ヘムへ渡り、さらに小胞内側ヘムへ渡る正規の経路の他に、細胞質側ヘムへ渡る別の経路やAsAから小胞内側ヘムへ直接渡る経路があることの反映と考えられる。酸化型標品をDEPC処理すると、細胞質側ヘムに配位したHis残基等が修飾され、AsAから細胞質側ヘムへの電子伝達が阻害される。DEPC処理時にAsAを共存させると、His残基の修飾は阻害され、Lys85残基のみが修飾される。この標品では、最も速いフェイズは、未処理標品の最も速いフェイズと比べると非常に遅いことからLys85残基はAsAとの結合において重要な役割をすると考えられる。また、低いpHでは未処理標品とAsAとの反応初期において顕著なラグが生じることがわかった。低いpHではシトクロムb561の基質結合部位に構造変化が起こり、b561-AsA複合体が生成され難くなる等の理由が考えられる[67]。 (h) 精製・可溶化したシトクロムb561を還元状態でリポソーム膜中に再構成すると、内腔中に封入したAsAに由来する膜貫通電子伝達反応が起こることがわかった。これは小胞外に添加した酸化型シトクロムcのヘムが還元されることから証明された。この還元反応は、前もって酸化型シトクロムb561をDEPC処理しておくと顕著に阻害されることがわかった。さらに小胞外に水溶性ドーパミンβ水酸化酵素を添加することによって、チラミン水酸化反応によるオクトパミンの生成反応をサポートできることもわかった。シトクロムb561はリポソーム膜中にクロマフィン小胞での向きとは逆に入っていることがトリプシン処理断片のSDS-PAGEとMALDI-TOF-MS解析の結果明らかとなった[68]。 (i) 我々はすでにシトクロムb561 1分子中には2つのヘムb中心が存在しており、完全酸化型では2種の異なったlow-spin種(gz=3.13とgz=3.69)がEPR測定により観測されることがわかっている。今回行った共鳴ラマン解析では酸化型、還元型ともに6配位型low-spin状態をとっていることがわかった。またEPR解析により、酸化型でDEPC処理したb561では細胞質側に位置すると思われるgz=3.69のヘム種がAsAでは還元できない形に変わることがわかった。しかし、小胞内側に位置すると思われるgz=3.13種については何の影響も無かった[69]。
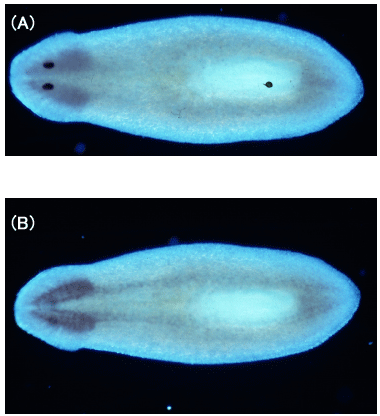
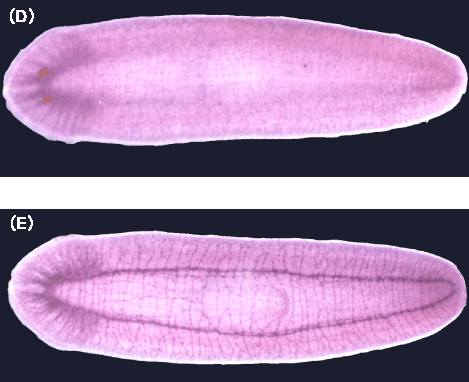
(6) 植物トウモロコシにおけるシトクロムb561のアスコルビン酸特異的電子伝達反応機構の解析 (a) 我々はこのトウモロコシ(Zea mays)由来のシトクロムb561遺伝子をクローニングし、アルコール資化性酵母Pichia pastorisを利用した大量発現系の構築に成功している。この発現系を利用することにより、トウモロコシそのものからの調製が殆ど不可能なシトクロムb561を大量に高純度に精製することが可能となった[77]。 (b) 酵母で発現させたトウモロコシシトクロムb561を高純度に精製する方法を確立した。b型ヘムを含み動物神経系型シトクロムb561と類似したスペクトルを示すこと、アスコルビン酸により容易に還元されること、等を明らかにした。さらにEPR法を用いた解析によって、動物神経系型シトクロムb561と同様に性質の異なる2種類のlow-spin型ヘムが存在することを明らかにした。 (c) 酸化型シトクロムb561へのアスコルビン酸(AsA)からの電子伝達反応をstopped-flow法により解析したところ、動物の神経内分泌シトクロムb561で見られたpH依存性と同じ振る舞いを示すことがわかった。AsAからの電子伝達には共通の分子機構が働いていることを示している。 (d) 現在、部位特異的変異体の作成とその大量精製にも成功し、それらの生化学的・生物物理学的解析を進めている。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||