

研究内容(トピックス)
クロマチン構造を介したDNA損傷認識の新たな制御機構を解明
- Kakumu, E., Nakanishi, S., Shiratori, H. M., Kato, A., Kobayashi, W., Machida, S., Yasuda, T., Adachi, N., Saito, N., Ikura, T., Kurumizaka, H., Kimura, H., Yokoi, M., Sakai, W., and Sugasawa, K. (2017) Xeroderma pigmentosum group C protein interacts with histones: regulation by acetylated states of histone H3. Genes Cells 22: 310-327.
ヒトのヌクレオチド除去修復は、DNA損傷に伴う二重らせん構造の歪みにより、正常な塩基対を形成できなくなった塩基をXPCタンパク質複合体が見つけて結合することで開始されます。また、紫外線で発生するDNA損傷に関しては、UV損傷DNA結合タンパク質(UV-DDB)が効率良く損傷を認識し、XPCを積極的に損傷部位に呼び込む働きがあることがわかっています。これらのDNA損傷認識因子がDNA損傷部位を認識するメカニズムについて分子レベルでの理解が進んできた一方、この過程が細胞内でどのように制御されているのかについては未だに良くわかっていません。特に、長大なゲノムDNAは細胞内でヒストンタンパク質に巻き付いてヌクレオソーム構造を形成しており、これがさらに何重にも折りたたまれて小さな核の中に収納されています。このような、いわゆるクロマチン構造はDNA結合タンパク質のDNAへのアクセスを阻害するものであり、ヒストンの翻訳後修飾(アセチル化、メチル化など)やクロマチンリモデリング因子などの働きによってクロマチン構造を適切に変換することが、特に遺伝子発現の制御では重要であることが明らかにされています。では、DNA損傷の認識でも共通のメカニズムが働いているのでしょうか?
私たちは、細胞に紫外線を照射してDNA損傷を与えた際、その周辺の領域でヒストンの脱アセチル化が誘導されている可能性を見出しました。ヒストン脱アセチル化酵素(HDAC)の阻害剤で細胞を処理すると、DNA損傷部位へのXPCの呼び込みや、それに続く損傷の修復が遅延することがわかりました。さらに、質量分析を用いた網羅的な解析から、XPCがヒストンH3およびヒストンH1と直接相互作用することを明らかにしました。特に、ヒストンH3に関しては、様々な翻訳後修飾の標的となるN末端テール領域を欠失したり、あらかじめヒストンアセチル化酵素CBPで処理してアセチル化修飾を導入することで、XPCとの相互作用が顕著に減弱することがわかりました。
以上の結果から、細胞内でDNA損傷が発生するとその周辺でヒストンの脱アセチル化を含むクロマチンの構造変化が誘導され、これによりXPCを積極的に呼び込んでいる可能性が考えられます。一般に、ヒストンの脱アセチル化はクロマチン構造の凝縮を引き起こし、転写因子のアクセスを阻害することで遺伝子発現を抑制するとされており、ヌクレオチド除去修復の開始段階ではクロマチン構造を介したまったく新しい制御機構が働いている可能性があります。DNA損傷部位周辺のクロマチン構造を凝縮させ、DNA損傷とXPCの局所的な濃度を高めることで、損傷を認識しやすくしているのかも知れません。
(本研究は、日本学術振興会科学研究費補助金・基盤研究(S)の支援により実施されました)
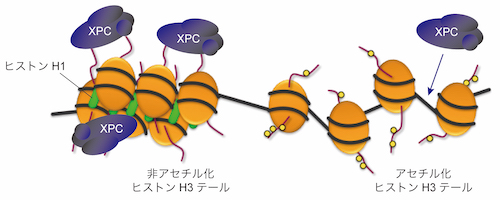
DNA損傷認識因子XPCの局在制御機構に関するモデル