

研究内容(トピックス)
DNA損傷認識を促進するヒストン脱アセチル化の役割を解明
- Kusakabe, M., Kakumu, E., Kurihara, F., Tsuchida, K., Maeda, T., Tada, H., Kusao, K., Kato, A., Yasuda, T., Matsuda, T., Nakao, M., Yokoi, M., Sakai, W., Sugasawa, K. (2022) Histone deacetylation regulates nucleotide excision repair through an interaction with the XPC protein. iScience 25: 104040.
ヌクレオチド除去修復(NER)は、紫外線や化学変異原など、主に環境由来の要因によって引き起こされる幅広いDNA損傷を取り除き、突然変異、さらにはがんなどの疾患の抑制に寄与している重要な生体防御機構です。ヒトのゲノム全体を対象とするNERは、色素性乾皮症C群(XPC)タンパク質を含む複合体(XPC-RAD23-CETN2ヘテロ三量体)が損傷部位を見つけて結合することによって開始されます。XPCがDNA損傷部位を認識するメカニズムについては、生化学・構造生物学的アプローチによって分子レベルでの理解が進んだ一方、この過程が細胞内でどのように制御されているのかについては不明な点が多く残されています。特に、長大なゲノムDNAの何処で、何時発生するかわからないDNA損傷を、XPCがどのようにして効率よく見つけ出しているのかは良くわかっていませんでした。
私たちはこれまでに、細胞に紫外線を照射してDNA損傷を与えた際、その周辺の領域で誘導されるヒストンの脱アセチル化がXPCの呼び込みを促進している可能性を報告しました(Kakumu et al., Genes Cells, 2017)。この詳細なメカニズムを解明するため、EGFPを融合したXPCを安定発現する細胞の核にレーザーを用いて局所紫外線刺激を行い、XPCのDNA損傷部位への呼び込みを経時的、定量的に解析しました。その結果、ヒストン脱アセチル化酵素HDAC1/2、及びHDAC1/2の活性化因子として知られるMTAタンパク質ファミリー(MTA1/2/3)の発現を抑制すると、DNA損傷部位へのXPCの呼び込みが減弱すること、さらにHDACとMTA自身がDNA損傷部位に呼び込まれることがわかりました。そこで、ラクトース・オペレーター(LacO)配列のアレイをゲノムの特定の位置に持つ細胞で、ラクトース・リプレッサー(LacR)とHDACの融合タンパク質を発現させ、LacO配列アレイの周辺で人為的にヒストンの脱アセチル化を誘導したところ、それだけでXPCをLacO配列アレイに呼び込めることが明らかになりました(図1)。これは、ヒストンの脱アセチル化がDNA損傷とは無関係にXPCの核内局在を制御できることをはじめて示した、画期的な成果と言えます。
XPCタンパク質の中央部分には、進化的にアミノ酸配列の保存性が低く、特定の立体構造を取らないと予想される、いわゆる天然変性領域が存在していますが、その機能は不明でした。私たちはこの中央領域が非アセチル化状態のヒストンH3のN末端テールと直接結合し、細胞内におけるXPCの局在制御に重要な役割を持つことを見出しました。XPC欠損細胞を親株として、中央領域を欠失した変異XPCタンパク質をEGFPと融合して発現させたところ、野生型XPCと比較してDNA損傷部位への呼び込みが減弱しており、また細胞のDNA修復活性の有意な低下が見られました。以上の結果から、1)ヒストンがアセチル化されたクロマチン領域で紫外線によってDNA損傷が発生すると、2)HDACやMTAを含む複合体が損傷部位に呼び込まれて周辺のヒストンを脱アセチル化し、3)ヒストンテールと中央領域との相互作用を介してXPCが損傷部位に呼び込まれやすくなり、4)局所的なXPCの濃度上昇によりDNA損傷の認識効率が上昇する、というモデルが考えられます(図2)。今後はこのモデルの検証を進めるとともに、HDACやMTAがDNA損傷部位へ呼び込まれるメカニズムを明らかにすることが重要な課題です。
(本研究は、日本学術振興会科学研究費補助金・基盤研究(S)の支援により実施されました)
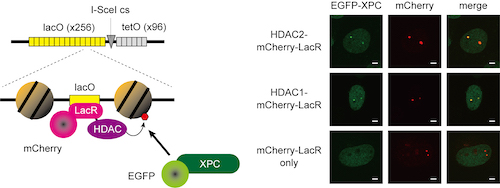
図1. LacO-LacRテザリングシステムを用いたXPCの局在制御解析
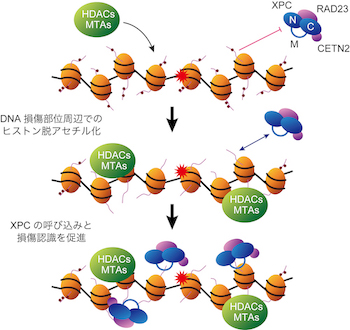
図2. ヒストンの脱アセチル化を介したDNA損傷認識制御機構のモデル