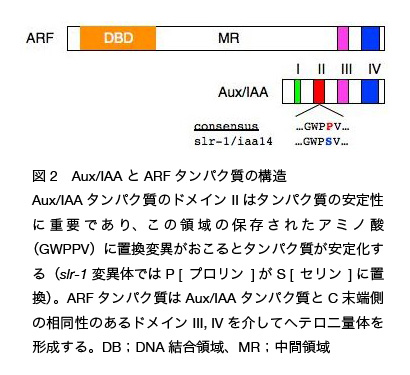
私たちはシロイヌナズナから側根を完全に欠失する変異体 solitary-root (slr) を単離しました(図 1)。slr 変異体は一遺伝子座の優性変異により、1)側根を全く形成しない、2)根毛をほとんど形成しない、3)根と胚軸の重力屈性が異常となる、4)オーキシンに対する感受性が低下する、などの多面的な表現型を引き起こします。slr のように側根を全く形成しない変異体はほとんど知られていなかったことから、この変異体の解析が側根形成開始機構の理解に有用であると考えられました。そこで側根形成の初期に発現するマーカー遺伝子(サイクリン B などの細胞周期関連遺伝子)を用いて詳しく解析すると、slr 変異が側根形成の初期段階つまり内鞘細胞の細胞分裂を抑えていることがわかり、SLR 遺伝子が側根原基の形成初期の細胞分裂・細胞分化に関わることが強く示唆されました。そこでマップベースクローニング法でこの原因遺伝子を突き止めると、SLR 遺伝子はオーキシン応答で働く転写抑制因子 Aux/IAA ファミリーの IAA14 をコードしており、slr 変異体では、IAA タンパク質の安定性に関わる保存領域(domain II)に 1アミノ酸置換を引き起こすミスセンス変異が生じていました(図 2)。シロイヌナズナには IAA タンパク質は 29 個存在しますが、いくつかの IAA タンパク質について domain II に同様なミスセンス変異が起こると、程度の差はありますがオーキシンを介した形態形成(側根形成、根毛形成、屈性、頂芽優勢、維管束分化、幼根形成など)に異常が生じます。slr 変異体でも、本来不安定な IAA14 タンパク質が変異によって安定化したために、オーキシン応答に異常が生じて側根形成が抑制されたと考えられます。ではどのようにして変異型 IAA14 タンパク質は側根形成を抑制したのでしょうか?
私たちはシロイヌナズナから側根を完全に欠失する変異体 solitary-root (slr) を単離しました(図 1)。slr 変異体は一遺伝子座の優性変異により、1)側根を全く形成しない、2)根毛をほとんど形成しない、3)根と胚軸の重力屈性が異常となる、4)オーキシンに対する感受性が低下する、などの多面的な表現型を引き起こします。slr のように側根を全く形成しない変異体はほとんど知られていなかったことから、この変異体の解析が側根形成開始機構の理解に有用であると考えられました。そこで側根形成の初期に発現するマーカー遺伝子(サイクリン B などの細胞周期関連遺伝子)を用いて詳しく解析すると、slr 変異が側根形成の初期段階つまり内鞘細胞の細胞分裂を抑えていることがわかり、SLR 遺伝子が側根原基の形成初期の細胞分裂・細胞分化に関わることが強く示唆されました。そこでマップベースクローニング法でこの原因遺伝子を突き止めると、SLR 遺伝子はオーキシン応答で働く転写抑制因子 Aux/IAA ファミリーの IAA14 をコードしており、slr 変異体では、IAA タンパク質の安定性に関わる保存領域(domain II)に 1アミノ酸置換を引き起こすミスセンス変異が生じていました(図 2)。シロイヌナズナには IAA タンパク質は 29 個存在しますが、いくつかの IAA タンパク質について domain II に同様なミスセンス変異が起こると、程度の差はありますがオーキシンを介した形態形成(側根形成、根毛形成、屈性、頂芽優勢、維管束分化、幼根形成など)に異常が生じます。slr 変異体でも、本来不安定な IAA14 タンパク質が変異によって安定化したために、オーキシン応答に異常が生じて側根形成が抑制されたと考えられます。ではどのようにして変異型 IAA14 タンパク質は側根形成を抑制したのでしょうか?この Aux/IAA ファミリーは ARF (AUXIN RESPONSE FACTOR) と呼ばれる別の転写因子ファミリーと相互作用を介してオーキシン応答性遺伝子群の転写を調節することが知られています(図 2)(相互作用はそれぞれのタンパク質の C 末側の良く似た領域で起こる)。一般にオーキシンによって制御される遺伝子のプロモーターにはオーキシン応答エレメント(AuxRE)と呼ばれるシス配列があり、ARF タンパク質はこのシス配列に結合します。ARF タンパク質はシロイヌナズナに 23 個ありますが、標的遺伝子の転写を活性化するものがいくつか知られています。一方 Aux/IAA タンパク質は ARF タンパク質と相互作用することによって ARF タンパク質の働きを抑制します。Aux/IAA タンパク質はもともと不安定なタンパク質ですが、オーキシンシグナルを介してさらにその分解が促進されることがわかっています(この分解は、オーキシン受容体 TIR1/AFBs を含む SCFTIR1/AFBs 複合体と 26S プロテアソームによって行われる)。すると相手の ARF タンパク質が機能するようになり、標的遺伝子の転写が活性化されます。おそらく野生型では内生のオーキシンシグナルによって SLR/IAA14 が分解されると相手の ARF が活性化し、側根形成を促進する遺伝子の転写を活性化すると考えられます。一方、slr 変異体では、本来不安定な IAA14 タンパク質が変異により安定化し、その結果、側根形成に関わる ARFs の機能が相互作用により恒常的に抑制され、側根形成促進に関わる遺伝子の転写が抑制されると考えられます。