home > Backgound 4. 側根形成を制御するARF7, ARF19
【オーキシンと側根形成】
4. 側根形成を制御する転写因子 ARF7、ARF19とその下流因子
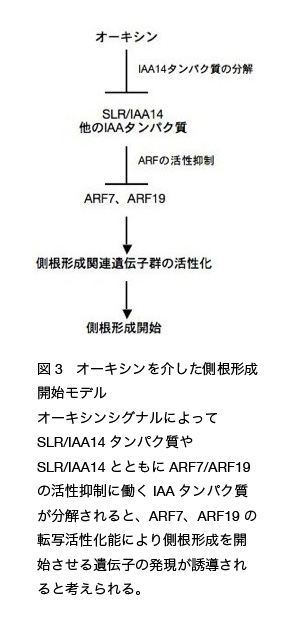
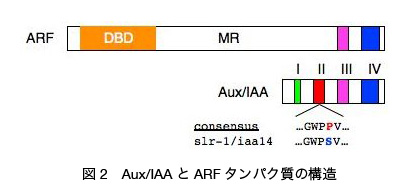
では SLR/IAA14 のパートナーとなる側根形成に関わる ARFs は存在するのでしょうか? ARF タンパク質群の機能を網羅的に解析する目的で、ARF 遺伝子群の単一および二重変異体の表現型について解析が行われました。その結果、米国カリフォルニア大学 Plant Gene Expression Center の Theologis 研究室などによる解析により、ARF7 と ARF19 の二重変異体 arf7 arf19 において単一変異体では見られない顕著な側根形成能の低下が観察されました(図 2)。この結果から ARF7 と ARF19 が側根形成促進に必要な ARFs として機能することが明らかとなりました。その後 Theologis 研究室と私たちのグループとの共同研究により、ARF7 と ARF19 が SLR/IAA14 と酵母細胞内で相互作用することや、ARF7 と ARF19 遺伝子が SLR/IAA14 遺伝子と同じ内鞘組織で働くことなどが確認され、オーキシンを介した側根形成開始に ARF7, ARF19, SLR/IAA14 を介した遺伝子発現調節が重要なことがわかっています(図 3)。
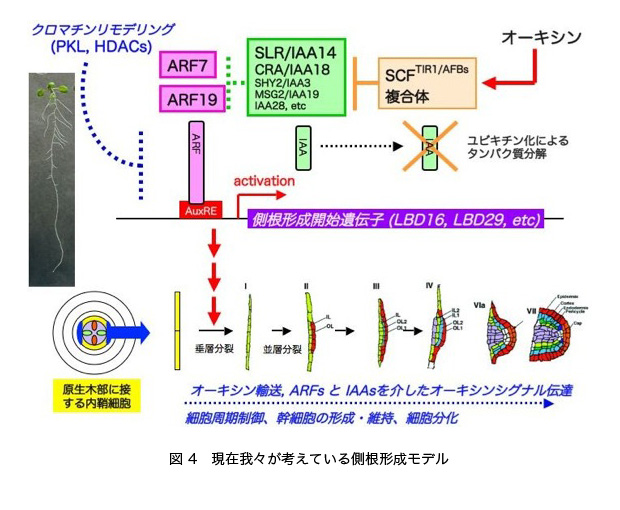
では、ARF7, ARF19, SLR/IAA14 は側根形成開始時にどのような遺伝子群を制御しているのでしょうか? そのような遺伝子を探索するために、野生型と slr 変異体、arf7 arf19 変異体におけるオーキシン存在下・非存在下における遺伝子発現プロファイルが解析されました。その結果、細胞周期、オーキシンホメオスタシス・オーキシン輸送・オーキシンシグナル伝達などに関与する遺伝子群の発現が変異体で顕著に低下していました。そして、最近私たちは、そのような遺伝子の中から側根形成で機能する ARF7, ARF19 の標的遺伝子として、LBD (Lateral Organ Boundaries-domain)/ASL (AS2-Like)ファミリーに属するLBD16/ASL18とLBD29/ASL16を見出しました(以下LBD16, LBD29と表記)。LBD16, LBD29は、オーキシンにより根で発現が誘導される遺伝子で、転写活性化因子をコードすると考えられています。また、これらのオーキシンによる発現誘導にはARF7, ARF19を必要とし、実際LBD16, LBD29のプロモーターにARF7, ARF19が結合することも確認しました。そして興味深いことに、これらのLBD16, LBD29タンパク質の過剰発現によってarf7 arf19二重変異体の側根形成能が部分的に回復しました。以上の結果から、ARF7, ARF19はLBD16, LBD29 を直接活性化することにより側根形成を制御することが明らかとなりました。
このLBD16は核局在タンパク質であり、転写抑制ドメインを付加したLBD16の過剰発現体において、側根形成が顕著に阻害されます。このことから、LBD16を含む複数のLBD/ASLタンパク質が転写活性化因子として重複した機能を持っており、ARF7, ARF19の下流で側根形成を正に制御していることが強く示唆されます。また、LBD/ASLタンパク質が、単子葉類イネでも冠根形成に働くことが示されました。このように近年、高等植物のオーキシン誘導性LBD/ASL遺伝子群の重要性が明らかになりつつあります。今後、これらの研究から、オーキシンを介した側根形成機構の理解が進むことが期待されます。