昆虫
昆虫は多様な生活様式で,陸上,地中そして水中や空中など様々な環境に適応し,地球上で最も繁栄している動物である.多くの昆虫は,寿命が短く,繁殖や世代交代にかかる時間が短い.このような短い寿命の中で,どのように環境に適応しているのだろうか.
適応的な行動の発現には,脳神経系が重要な役割を担っている.動物は,周囲の環境から様々な信号を刺激として感覚器で受容する.受容した感覚信号は,中枢神経系で処理,統合されて,必要な情報が抽出される.そして,記憶や生理状態と照合されて運動制御信号が生成され,効果器に出力されて運動や行動が発現する.
動物の適応的な行動発現を司る神経生理機構を理解することは,神経生物学研究における重要な課題である.昆虫の脳神経系は,10万から100万個程度の神経細胞から構成され,ヒトの脳を構成する神経細胞の数と比較すると,わずか100万分の1程度であり,小規模な神経系で構成されている.このような小規模な脳神経系で,昆虫が脊椎動物の行動にもひけをとらないほどの運動や行動パタンの多様性を表出するのは,本能的な行動の他にも,経験にもとづく適応行動の発現機構(学習と記憶の仕組み)を備えているからである.
超高速度運動の制御

脅威に対する行動選択


オキナワアギトアリの腹部を筆の先端背触ると,前方へ突進する回避行動を示すときと,刺激源に対して威嚇行動を示す場合がある.回避行動を示す個体と威嚇行動を示す硬いの脳内の生体アミン量を比較すると,威嚇行動を示す個体の方が有意にセロトニン,オクトパミン,ドーパミンの量が多いことがわかった.脳内の生体アミン類は,神経伝達物質や神経修飾物質,神経ホルモンとして機能し,個体の生理状態(内部状態)を変化させる.
闘争行動

秋の夜長に耳にするコオロギの鳴き声(発音)を方も多いことだろう.この鳴き声は,雄が交尾相手の雌を呼び寄せる誘引歌である.ここで紹介するコオロギは,クロコオロギと呼ばれ,沖縄県の八重山諸島に棲息するコオロギで,土中の穴を住処とする.雄コオロギは,同種の雄が巣穴付近の縄張りに侵入すると激しく威嚇や攻撃を始め,闘争が起こる.コオロギの闘争は,中国では1000年以上昔から闘蟋と呼ばれる賭け事として親しまれている.
コオロギは,接近してきた相手の体表を触角で触り,体表を覆う炭化水素の化合物を手掛かりに相手の性別や加齢の状態を識別する.この物質は体表フェロモンとして働き,雌のものであれば求愛行動を,雄のものであれば威嚇や攻撃行動を雄コオロギに誘発する.闘争は数秒から数十秒で終結し,勝敗結果は,他の動物と同様に,2個体間の優劣関係(社会的順位)の形成に重要である.また敗者は,闘争終結後3時間以上,個体によっては1日以上もの間,攻撃性が減退し,他の雄個体を忌避するようになる.
コオロギの敗者で見られる「攻撃性の減退状態が持続する」脳の仕組みと,ヒトの社交障害や発達障害等の発生メカニズムには類似する点はないだろうか.攻撃性や暴力は,ヒト社会でも取り上げられる重要な課題であり,また,殆どの動物に共通して備わる行動である.攻撃性の生理学的なメカニズムや進化的な意義については未解明な点が多く,昆虫の攻撃性や闘争の発現メカニズムを解明することで,その理解や,更には社交障害や発達障害などの発生メカニズムの解明にもつながる.
脳の設計を理解するためには脳を構成する細胞の分子基盤を理解することが不可欠です.しかしながら,分子レベルの研究から得られた結果をそのまま個体行動の理解へと結びつけるには大きな飛躍があります.神経細胞や神経回路の生理学を理解し,ボトムアップ的に知見を積み上げて総合的に理解する作業が必要です.脳には一酸化窒素,生体アミン類など,神経伝達物質・神経修飾物質として働く生理活性物質があります.これらの物質の機能的な役割を理解するため,分子生物学的な方法で機能解析を進めています.
コオロギは,行動学的な知見とその基盤となる神経系の生理機能と個々の細胞の生理学的・形態学的な知見が豊富な実験動物である.私たちの研究室では,遺伝子改変,RNAiなどの方法で神経系の動作修飾することで,闘争行動の発現基盤となる神経生理機構,分子機構の解明にも取り組んでいます.
脳における一酸化窒素 (NO) シグナル伝達系
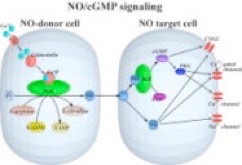
これまでの神経系における NO の生理機能・作用機序の研究は,脊椎動物を中心に研究が進められており,NO が学習・記憶などの脳の高次機能に関わることが報告されています.一方,我々の研究室で扱うコオロギなどの無脊椎動物の神経系においても,NO が広範な脳領野で生産され神経細胞間の情報伝達を担うことがわかっていますが,無脊椎動物脳における NO の生理機能についてはほとんど研究が進められていません.我々の研究室では,コオロギなどの昆虫の神経系を材料に,NO による攻撃行動の調節機構に焦点を当てて研究を進めています.
NO は可溶性グアニル酸シクラーゼ (soluble gualylyl cyclase; sGC) を活性化し,標的細胞の細胞内 cGMP 濃度を上昇させます.cGMP は細胞内でセカンドメッセンジャーとしてはたらき,cGMP 依存性タンパク質リン酸化酵素 (protein kinase G; PKG) や,細胞膜上の環状ヌクレオチド依存性イオンチャネル (cyclic nucleotide-gated cation channel; CNG channel) を活性化します.このような下流のシグナル伝達系を介して,NO は標的細胞における短期的・中期的な細胞機能の可塑的変化を引き起こします.
我々は NO 標的細胞における NO 下流の情報伝達系に着目し,現在,コオロギ脳で発現する NO 下流シグナル伝達系関連因子群 (sGC, PKG, CNG channel 遺伝子) を同定し,それらの遺伝子群の発現解析をおこなっています.また,組織学的手法によって NO 産生細胞や NO の標的細胞の脳内分布を調査しています.将来的には,現在の研究から得られた知見を遺伝子導入実験と組み合わせ NO 下流のシグナル伝達系の機能修飾実験などをおこなうことで,NO による攻撃行動の調節機構を分子レベル・細胞レベルから明らかにしていきたいと考えています.
モデル構築とロボット実験

昆虫とほ乳類の脳内物質の機能的類似性: 神経系における一酸化窒素(NO)の働き
NOは,ヘモグロビンとの親和性が酸素と比べ24万倍以上も高いことから,生体にとっては有害な物質と考えられがちであるが,生体の様々な器官や組織には,NOを合成する酵素(NOS)が局在し,細胞間の情報伝達を担う拡散性の生理活性分子として働いている.
昆虫の脳は,ヒトの脳に比べて非常に小さく,神経系を構成する細胞数は100万個程度と言われている.ヒトの脳がおよそ1000億個の細胞から構成されている(Hubel,1979)ことを考えると,昆虫の優れた環境適応能に驚かされる.昆虫脳は,小規模なゆえに,脳を構成する神経細胞が生理学的にも形態学的にも同定可能で,細胞の機能と表出する行動を関連づけて研究が行われてきた.
柔軟で可塑的な行動の発現には,脳内で情報処理や調節にかかわる神経作用物質の役割が重要である.中でも生体アミン類は,複雑な行動の発現や修飾にかかわり,ヒトを含めた脊椎動物から昆虫などの無脊椎動物の神経系で主要な役割を担っている.また,生体アミンは昆虫と哺乳類で非常に類似した機能を担うことが示されている.
脳におけるセロトニン(5-HT)やドーパミン(DA)神経回路における神経修飾の機能障害は,統合失調症や鬱などの発症とも関係が深い.また,ほ乳類ではエピネフリン(E)や5-HTそして,一酸化窒素(NO)などの物質も攻撃性の調節にかかわる.昆虫でもNOや5-HTそして,OAやチラミン(TA)などが攻撃性の調節に関わる物質として知られている.昆虫の脳で働くOAやTAは,哺乳類におけるEやノルエピネフリン(NE)と機能的に類似な働きを担うと考えられている.哺乳類と昆虫は異なる集中神経系を進化させて地球環境に適応してきた.しかし,OA/TAとE/NEの機能的類似性が示唆するように,神経系をひとつのシステムとして考えれば,コオロギの闘争行動の発現メカニズムの中にはヒトの発達障害や社交障害の発生メカニズムに類似した神経制御メカニズムが見えてくる.
学習と記憶にかかわるNO
NOはおよそ100μm/秒の速度で三次元的に拡散し,標的細胞の細胞膜を自由に透過できる.この様な性質から,NOは順行性の伝達物質としてばかりではなく,逆行性の伝達物質としても働き,シナプスを形成していない細胞間のコミュニケーションを担うことが可能である.NOは神経可塑性を作る役割を担っている物質のひとつと考えられ,コオロギでも,NOシグナル系が長期記憶の形成過程で重要な働きをすることがわかっている.
闘争行動にかかわるNO
オスのコオロギは別のオスに出会うと,激しい噛みつき合いの闘争を始める.闘争に負けたオスは,その後,1時間以上他のオスに対して攻撃を仕掛けずに忌避行動をするようになる.この闘争の勝敗に依存した行動選択には,脳内のNOが関与する.
