|
|
「独創性とは何か。万人の目の前にありながらも、まだ名前を持たず、まだ呼ばれたことのないものを見ることである」
フリードリッヒ・ニーチェ
園芸家は当時も今も、多くの観賞植物で新しい色彩や形態を持つ変種・品種を作ることに熱心である。メンデルもこうした「変わりもの(sports)」の存在をよく知っていたし、同じ種内あるいは異なる種間で交配(交雑)を行う度に同じような雑種型が生じるという先人達が観察していた規則性についても知っていた。メンデルは、そうした雑種の子孫では一体どんなものがどのような規則性を持って現れてくるかに興味を持った。新しいものの存在や現象に初めて気づいたのではない。古くから知られていたものに科学の眼を向け、明確な意図をもって、綿密な計画と用意周到な準備のもとに多数の交配実験を行い、その後代の子孫を詳細に観察し、結果を数学的に解析するなど独創的な方法でそこに潜む真理を初めて明らかにした。メンデルはまさに遺伝学の魁であった。
メンデルの原著論文
 図1 メンデルがエンドウを育てた畑
図1 メンデルがエンドウを育てた畑
|
 図2 メンデルのエンドウ
図2 メンデルのエンドウ
|
1854年、メンデルは、聖トーマス大修道院の裏庭にナップ院長が用意してくれた縦35メートル、横3メートルの小さな畑(図1)と温室を実験場として、ジョセフ・マレシュという従僕一人とともにエンドウ(図2)を相手に植物雑種の研究を始めた。2年間の予備実験と用意周到に計画した8年間に及ぶ交配実験から得た膨大なデータの綿密な数量的解析を経て、1865年2月8日と3月8日の2回、ブルノ博物学協会の例会で実験結果を発表し、翌1866年に同会の雑誌に「植物雑種の実験」という題目のドイツ語論文として公表した。この原著論文は、1901年にイギリスのウィリアム・ベートソンによって英語に翻訳され、ヨーロッパの学術界に紹介され、その後に数回の改訂を経て1996年に電子版として公開されている(Druery, C.T. and Bateson, W. (1901) Experiments in plant hybridization. Journal of the Royal Horticultural Society 26: 1-32. 1996, ©Electronic Scholarly Publishing Project. ; 山下孝介(訳編)(1972)メンデリズムの基礎 ―メンデルの「植物雑種に関する実験」ほかー 裳華房; 「雑種植物の研究」グレゴール・メンデル(著)岩槻邦男・須原凖平(翻訳)(1999)岩波文庫 岩波書店)。本章では、「植物雑種の実験」に書かれた内容のうち論文の理解に必要不可欠と思われる項目について順次見ていくことにする。なお、論文の序論(1章)、エンドウ以外の植物雑種を用いた実験(10章)と結語(11章)については、2016年にイギリス科学史協会から出版された最新の英語翻訳論文とその解説論文を参照した。(「植物雑種の実験」に関する最新の翻訳版および解説書は以下である。(Mendel, G. (2016) Experiments on Plant Hybrids (1866). Translation by Müller-Wille, S. and Hall, K. British Society for the History of Science Translation Series. ; Abbott, S. and Fairbanks, D.J. (2016) Experiments on plant hybrids by Gregor Mendel. Genetics 204: 407-422.)。
メンデルの「植物雑種の実験」は、遺伝に関する明確な科学的概念が未だなく、遺伝現象を記述するための適切な術語も存在しない時代に書かれた。論文は“形質の子孫への伝達を支配する遺伝法則”の発見に至るメンデルの意図と思考を忠実に辿って書かれており、その内容は詳細かつ正確である。しかし、今日の科学論文では必須な引用論文の記載やデータを分かり易く示した図表がない。本章では、ベイトソンの英語翻訳文に基づき、現在の学術用語でいう形質 (trait) は「形態 (form) あるいは性質 (character) 」、遺伝子 (gene) は「因子 (factor) 」、対立遺伝子 (allele) は「対照的な形態あるいは性質」などの述語をそのまま用いた(遺伝を扱う学問を遺伝学「genetics」と名付けたのはウィリアム・ベイトソンで、1906年にロンドンで開催された植物雑種に関する国際会議の場であった。メンデルが原著論文で用いた性質あるいは形態に形質という用語を与え、対照的な形態あるいは性質を決める因子を対立遺伝子と名付け、形質に関して個体が表す表現型 (phenotype) と個体が保有する遺伝子型 (genotype) を区別し、さらに同じ二つの対立遺伝子をもつ個体の遺伝子型をホモ接合体 (homozygote) 、二つの対立遺伝子が異なる個体の遺伝子型をヘテロ接合体 (heterozygote) としたのもベイトソンであった。一方、メンデルの因子を遺伝子「gene」と名付けたのはウィリアム・ヨハンセン (1909年) である)。なおメンデルは、両親と雑種(hybrid)および雑種の自家受粉で得た後代が表す「形態あるいは性質」と卵細胞と花粉細胞を通じて伝達された「因子」の組み合わせの両方をA, B, Cなどのイタリック体(斜体)の文字記号で表記しており、親と同一の表現型を子に与える遺伝子型ホモ接合の個体を安定親型と呼び、こうした個体はAあるいはaなどの1文字で表し、ヘテロ接合の雑種型個体をAaなどの2文字で表しているが、ここではそのままとした。遺伝を担う因子と形態・性質の厳密な区別および単数性(半数性)の配偶子と2倍性の接合体の区別を正確に認識していたメンデルにあってなお両者の表記が曖昧なままであった事実は、遺伝の概念がなかった時代にこれを記述・説明せざるを得なかったメンデルの苦労を忍ばせるものであろう。なお、メンデルの主張を記述した文章は鉤括弧で囲み、メンデルが重要として斜体文字で記述した部分はそのまま斜体として残した。
1章)序論
序論では、メンデルが研究を手がけた動機、意図・目的と意義が述べられている。メンデルはまず、「花色の異なる新しい変異を備えた観賞用の園芸植物を手に入れようとして過去に行った人為交配実験から得ていた結果に触発されて、実験を始めた」と語っている。メンデルは述べていないが、これらのうちにはカーネーションや特に好きだったフクシアが含まれていた。続いて、すでに報告されていた他の研究者の実験内容にも触れて、「同一種間の交配では、雑種がいつも同じ性質を示す驚くほどの法則性に興味を引かれたことから、雑種の後代ではどのような子孫がどのような割合で現れるかを追跡調査する目的で実験を行った」と説明している。
メンデルは、先行する研究者であったヨーゼフ・ケールロイター、カール・ゲルトナー、マックス・ヴィヒューラの他に2名の名を挙げて、彼らが辛抱強く生涯の一部を捧げて得た観察結果、特にゲルトナーが著書「植物界における雑種の形成」で、ヴィヒューラが著書「ヤナギ類の雑種について明らかにされた植物界における雑種の受精」で記録した価値ある観察結果がこの問題に与えた貢献に敬意を払い、当時の現状を次のように分析している。これらの研究では、「雑種の形成と子孫の発達を支配する一般的な法則が依然として明らかではない」、「雑種の子孫で出現する異なる形態を示す個体の数を決定し、それらを個別の世代で正確に分類し、さらにそれらの間の統計的な関係を確証するような実験が未だなされていない」、「そのような広範な実験を実施するためには多大な労力が要求されるだろうが、そうしてこそ、生物の進化史上で軽視することができないほどの重要な意味を持つ問題の解決に至ることが可能となるだろう」と述べている。その上で、自身が企てここに論文とした雑種の研究は、「比較的小さなグループの植物を対象とした限定的なものではあるが、前例のないほどに詳細で完璧な計画に沿って、8年もの歳月をかけて実施した実験であり、過小評価すべきでない価値をもつ望ましい結果をもたらしたものである」と控えめだが誇らしく述べ、「読者の自由な議論と批判に委ねたい」との希望で序論を締めくくっている。
2章)実験材料の選択
この章でメンデルは、「すべての実験の価値と有効性は、そのために使われた材料が適切であるかどうか、またそれを目的に適ったやり方で使ったかどうかにかかっている」として、実験材料と方法の重要性を強調している。メンデルの指摘は以下の通りである。「実験計画で用いる親植物が備えるべき二つの特徴、すなわち、一定あるいは安定した識別可能な形態や性質を示すこと、雑種の開花期に他植物体の花粉による予期しない受粉が起こらないこと、または人為的な操作でそれを避けることが可能であること、さらに、雑種とその後代の稔性(種子の着粒率)に悪影響がないこと、雑種どうしの関係と雑種と親植物との関係を明らかにするために得られた子孫は例外なく全て評価の対象とすること」。こうした条件を勘案した上で、メンデルは、特有な花の構造を示すマメ科(Leguminosae)植物に着目し、エンドウ属のエンドウ(Pisum sativum L.)を最も適した実験材料として選んだ。
続いてメンデルは、エンドウの特徴を以下の様に詳細に分析し、「安定で容易かつ確実に識別できる形態あるいは性質を備えた系統があること、雑種の稔性が親植物と同等に正常であること、雌蕊と雄蕊が蝶型の花の下方につく2枚の竜骨弁の内部に閉じ込められていて、雌蕊による雄蕊からの花粉の受け取り(受粉)が開花の前に行われほぼ完全な自殖稔性を持つこと、面倒ではあるが開花前の花から竜骨弁を除去し、虫眼鏡を使ってピンセットで雄蕊を除去した後に、別の系統・個体の花粉を刷毛で母(種子親)植物の花柱に塗布することで人為交配が可能であること、圃場と温室のどちらでも栽培が容易であること、世代時間が比較的短いこと、一回の受粉で比較的多くの種子が採れること」を、エンドウを実験材料に選んだ根拠として挙げている。実際にメンデルは種子商人から購入した34品種・系統を予備実験として2年間かけて育てて観察した後、上記の条件に適った22品種・系統を選抜し、以後の交配実験に用いた。メンデルは選んだ系統の分類学上の位置づけについても専門家の見解を紹介し、これを自身の選択の正当性の根拠として主張している。メンデルが選んだ22品種・系統には、品種と亜種だけでなく近縁の別種が3つ含まれていたが、メンデルはこれらの区別は、事実上困難であるうえに、問題とする実験の目的にとって、さして重要ではないと述べている。当時は、品種や亜種の識別は困難であり、近縁種との明確な区別は事実上不可能であった。ダーウィンも、「種の起原」の中で、「すべての博物学者を納得させられる種の定義はまだ一つもない。それでも博物学者は誰もが、種を語るときには自分が意味するものを漠然と知っている」と語っている(レベッカ・ステフォフ著 西田美緒子訳(2008)「オックスフォド 科学の肖像 ダーウィン:世界を揺るがした進化の革命」 オーウェン・ギンガリッチ編集代表、大月書店)。
3章)実験の区分とその配置
メンデルは、「ひとつあるいは複数の性質について安定して相違する親植物を交配で組み合わせると、親植物が共通に持つ性質は変わることなく雑種とその子孫に安定して伝えられるが、親植物で対照的な形で相違する性質はそれぞれ雑種で合一し、その子孫で様々に変化して現れる」と云う、先人による多くの実験で明らかになっていた観察結果をまず述べる。続いて、研究者たちが過去の実験から得たこうした観察結果を引用して、明瞭な対の形で相違する性質が示すそのような変化が後代で再び現れるという現象のうちにある法則性を見いだすことが実験の目的であるから、実施する実験は「両親がもつ安定して相違する性質の数と同じだけの個別実験で実施する必要がある」と指摘している。
メンデルは、茎の長さ(草丈)と色、葉の大きさと形、花のつき方と色と大きさ、花梗の長さ、莢の色と形と大きさ、種子の形と大きさ、種皮の色とアルブミンの色すなわち子葉の色からなる15の異なる性質を調査し、識別が誤りなく明瞭で、目的に最も叶った7つの性質を最終的に選び出した(子葉の色をメンデルは種子のアルブミンの色と呼んだ。この性質は、双子葉植物であるエンドウの種子の主要部分を占め、次代の個体に発達する子葉の性質を指すので、以後はアルブミンの色は子葉の色と記述することにする)。なお、以下に示すメンデルが選んだ7つの性質のうち第1と第2の性質は種子の性質だから、雑種個体につく種子の観察で直接に次代の判定が可能だが、その他の性質は雑種種子を育てた雑種植物体でのみ評価が可能で判定に時間がかかった。ここで、種子に関する2つの形質を簡単に説明しておく。当時、胚乳形質は母親に由来する形質であるとされていたが、メンデルの結果は、この見解が誤りであることを示していた。花粉親の形質が種子の胚乳に現れる現象を特にキセニアといい、トウモロコシの胚乳の色(黄色と白色)がよく知られた例である。胚乳形質が母親形質であると仮定すると、この一見不思議に思われる現象は1881年にヴィルヘルム・F.フォッケにより初めて記載され、1898年にロシアのセルゲイ・G. ナヴァシンによるマルタユリの重複受精の発見でその仕組みが明らかになった。実は、キセニアと呼ばれる農業上重要なこの現象も、両性が関与する通常の形質と同様に雌雄配偶子の融合の結果なのである。重複受精とは、被子植物で見られる受精様式で、花粉管の先端にある二つの精細胞(雄性配偶子)による受精が胚嚢中の卵細胞と中心細胞の2カ所で行われる現象であった。1900年にドイツ人エードゥアルト・A シュトラースブルガーは、卵細胞にある卵核と精核との受精を胚形成のための生殖受精と呼び、中心細胞の極核と精核との受精を胚乳形成のための栄養受精と呼んだ。nを生殖細胞のもつ染色体数とすれば、生殖受精によって生じる胚の染色体数は2nとなり、栄養受精によって生じる胚乳の染色体数は3nとなる。第3の性質は種皮の色と花の色の二つに関する性質で、子孫では常に同一の組み合わせで発現する。これと第7までの4つの性質はすべて植物体を育てて初めて決定可能な性質であった。メンデルはこれら7つの性質を評価する際に注意すべき点を、以下のように丁寧に記載し説明している。
1.完熟種子の形態:表面が滑らかで丸い種子(丸種子)とシワ状となる種子(シワ種子)。丸種子でもシワの形状が時折観察されるが、そうしたシワは表面のみに表れるのに対して、シワ種子は、近縁種のP. quadratumが示すような、不規則に角張った種子の内部にまで及ぶ深いシワを安定して表す。
2.子葉の色:黄色の子葉(黄色種子)と緑色の子葉(緑色種子)。黄色種子の実際の色は、うすい黄色か鮮やかな黄色あるいはオレンジ色で色調には一定の幅が見られるが、緑色種子はどれも濃い緑色である。種皮は透明だから、内部にある子葉の色は外部からの観察で容易に識別が可能である。
3.種皮の色と花の色:すみれ色の種皮ですみれ色の花(この性質はすみれ色の花と呼ぶ)と白色あるいは灰色の種皮で白色の花(白色の花と呼ぶ)。すみれ色の種皮には、灰色あるいは灰褐色か皮革のように褐色ですみれ色の斑点があったりなかったりするが、基本的に種皮はすみれ色である。すみれ色の種皮をもつ種子から育てた植物体は紫色の側方翼弁のあるすみれ色の花をつけ、茎の葉腋に赤みがかった斑点が生じる。灰色・白色の種皮をもつ種子から育てた植物体の花は全て白色であるが、種子は煮ると暗褐色に変色する。
4.完熟した莢の形:膨らんだ莢とくびれた莢。膨らんだ莢は莢全体が膨らんでおり、一方、くびれた莢は、P. saccharatumの性質と同じで、内部の種子の位置を反映して表面に表れるくびれが明瞭である。
5.未熟な莢の色:緑色の莢と黄色の莢。緑色の莢は実際には薄い緑色か濃い緑色で、黄色の莢は鮮やかな黄色を示し、茎、葉脈と萼も同様に鮮黄色である。
6.花のつき方:茎(主軸)に沿って花が散在してつく腋性花序と茎の先端に花がつく頂性花序。頂性花序では、P.umbellatumのように、茎の上部に花が笠のような形につく。
7.茎の長さ:茎が長く背丈が高い性質(長い茎と呼ぶ)と茎が短く背丈が低い性質(短い茎と呼ぶ)。判定を確かにするために、長い茎では長軸の長さが6~7フィートのものを、短い茎では0.75~1.5フィートのものを実験に供した。
続いてメンデルは交配実験の区分、その配置と植物個体の栽培条件について述べているが、重要な関連事項は結果の項に回して、ここではメンデルが交配に用いた個体数とともに、強調した以下の注意事項を見てみよう。メンデルは、性質1から7について、それぞれ15、10、10、10、5、10、10個体を母個体に用いて実験を始めた。さらに、雑種を作る際には、多くの個体から生育の旺盛なものだけを交配親に選んだ。生育の良くない個体では、後代で開花が見られず結実が不良なものが出現し、予期しない誤った結果を得る可能性が高かったからである。また、望まない受粉を避けるために、昆虫、特に竜骨弁を押し開いて花の内部に卵を産みつけるエンドウゾウムシには特別の注意を払った。加えて、両親の貢献度を比較するために、全ての組み合わせで母親(種子親)と父親(花粉親)を交互に換えた正逆交雑を実施している。
次章からは、いよいよ実験結果である。メンデルは各章のはじめにまず実験で得られた結果を取りまとめ、その後にそれぞれ結論的な解説を加えている。
4章)雑種(F1世代)の性質・形態
メンデルは、先に鑑賞用の園芸植物で自ら行った実験から、「葉の形や大きさ、植物体各部の毛の有無など少数の性質については、雑種が親の中間型となる場合が認められるが、多くの性質については、両親のどちらか一方の性質が著しい優位を占め、他方が認められないことが多い」という観察結果を既に得ていたが、エンドウで行った交配実験の結果はまさにそれを確認するものであった。
調査した7対の性質のそれぞれについて異なる両親間の雑種第一代F1は常にどちらか一方の親と区別できないほどに似ていた。ここでメンデルは、雑種への伝達が完全でほとんど変化が見られなかった親の性質、すなわち雑種で表れた性質を顕性(優性)、雑種で表れなかった性質を潜性(劣性)と名付けた(メンデルは雑種で現れない性質を「休む」あるいは「引き下がる」を意味するrecessiveと名付けたのであって、「劣る」を意味する劣性という日本語の述語は、誤解を招きかねず、相応しくない。メンデルの「植物雑種の実験」の抄訳・解説本「メンデリズムの基礎」を著した山下孝介は、優性、劣性ではなく、顕性、潜性という用語が適切であろうと述べている。実際に中国では顕性、潜性が用いられている。なお、日本遺伝学会では、議論の末、2017年9月に出版した[遺伝単: 遺伝学用語集 対訳付き、『生物の科学 遺伝』別冊N0.22)]で、優性の代わりに顕性を劣性の代わりに潜性を用いることを推奨している。本書では以後、これに習い顕性・潜性と云う述語を用いることにする)。
7対の性質について雑種で表れた顕性の性質は、丸種子、黄色種子、すみれ色の花、膨らんだ莢、緑色の莢、腋生および長い茎で、一方、雑種で表れなかった潜性の性質は、それぞれシワ種子、緑色種子、白色の花、くびれた莢、黄色の莢、頂生および短い茎だった。雑種では両親のどちらか一方の性質のみが表れるこの現象は後に顕性の法則(メンデルの第三法則)と呼ばれることになる。なお、最後の性質(茎の長さ)について、メンデルは、雑種が顕性の親よりも大きな値を示すこと、すなわち現在では農業上重要な雑種強勢(ヘテロシス)として知られる現象にも気がつき、これについて注意を促している。
5章)雑種の第二代(F2世代)
メンデルは、まず以下の決定的に重要な結果を述べる。すなわち、調査した7対の性質の全てについて、例外なく、雑種のF1世代を自殖(自家受精)した雑種第二代のF2世代では、顕性の性質とともに潜性形質が再び出現し、しかも顕性の性質を表す個体と潜性の性質を表す個体の出現頻度は常に3:1の割合であることを明らかにする。
なお、メンデルの論文では雑種世代の表し方について、現在の表記法とは異なり誤解を招く表現が用いられている。すなわち、メンデルは雑種当代(第一代)のF1を雑種と呼び、F1の自家受粉で得られた雑種第二代のF2を雑種から導かれた雑種第一代と呼んでいるが、混乱を避けるために、以下では章題も含めて、現在の表記法に従って、それぞれ雑種第一代F1あるいは単にF1、雑種第二代F2あるいは単にF2と表示することにする。
以下は具体的な実験結果である。なお、正逆交配(種子親と花粉親を入れ替えた交配)で得た雑種でも全く同じ結果であったから、メンデルは両者をまとめて数量的に取り扱っている(図3)。
実験1)種子の形態について
実験2年目に253個体の雑種第一代F1から、自殖により7,324個の雑種第二代F2種子を得た。そのうち丸種子が5,474個、角張ったシワ種子が1,850個で、両者の比率は2.96対1であった。
 図3 メンデルが交配に用いたエンドウの7つの変わりものとF2世代の分離
図3 メンデルが交配に用いたエンドウの7つの変わりものとF2世代の分離
|
実験2)種子の子葉の色について
実験1と同様に、258個体のF1から8,023個のF2種子を得た。そのうち黄色種子が6,022個、緑色種子が2,001個で、両者の比率は3.01対1であった。
実験1では母親として用いた15個体の雑種個体のうち10個体について、実験2では10個体すべての母親雑種個体について、個体ごとに生データを示している。示された生データでは、実験1では顕性:潜性を表したF2種子の観測数が19 : 10 (1.9 : 1)から28 : 6(4.7 : 1)まで振れており、実験2では同じく24:13(1.8 : 1)から44:9(4.9 : 1)であった。上に挙げたF2世代の分離比は、得られた結果をそれぞれ実験ごとに取りまとめた合計と平均値である。
実験3で調査した花の色、実験4の莢の形、実験5の未熟な莢の色、実験6の花のつき方および実験7の茎の長さに関する結果はそれぞれ、3.15:1(全観察数929個体)、2.95:1(1,181個体)、2.82:1(580個体)、3.14:1(858個体)、2.84:1(1,064個体)で、実験結果を全体として集計すると、優性が2.98で劣性が1、すなわちF2全体の分離比は3:1の近似であった。なお、メンデルは両親のどちらか一方の性質に移りつつあるような中間体は全く観察できなかったと述べている。
メンデルは以上の結果から、顕性は、顕性親が示す性質と雑種の性質という二つの違った意味を併せもつという推論に到達する。すなわち、顕性を示すF2個体は、安定な顕性親型と雑種型の2つのグループからなり、どちらのグループかは次代を調査して初めて分かる。すなわち顕生親型は次代に親がもつ顕性の性質のみを伝えるが、一方、雑種型ではF2世代と同様にF3世代でも顕性と潜性が3:1の比で再び分離するはずであると推論して、これを明らかにする目的で、以下の実験に進んでいる。
6章) 雑種の第三世代(F3世代)
メンデルの予想通り、雑種第一世代F1では表れず、第二世代F2で潜性親の性質を再び表した個体は次の第三世代F3でも変化せず潜性であった。F2で顕性を示した個体では状況が異なり、その3分の2ではF2と同様にF3で再び顕性と潜性が3:1の割合で生じた。すなわち3分の2の顕性個体は雑種型の振る舞いを示したが、残りの3分の1は顕性親の性質のみを示す安定親型であった。以下に、実際の実験結果を示す。
実験1)種子の形について
F2の丸種子から565のF2個体を得た。F2個体に着粒したF3種子から育てたF3個体のうち3分の1の193個体は丸種子のみを着粒する安定親型であった。一方、残り3分の2の372個体はF3で丸種子とシワ種子を3:1の割合で着粒する雑種型であった。このように、F2世代で顕性を示した個体のうち、雑種型個体と安定親型個体の割合は1.93:1だった。
実験2)種子の子葉の色について
F2の黄色種子から519のF3個体を得た。そのうち、3分の1の166個体はF3で黄色種子のみを着粒する安定親型を示し、残り3分の2の353個体はF3で黄色種子と緑色種子を3:1の割合で着粒する雑種型であった。従って、F2世代で顕性を示した個体のうち、雑種型個体と安定親型個体の割合は2.13:1だった。
他の5対の性質3から7につては、F2世代で顕性を示したそれぞれ100個体を選び、各F2個体から得たそれぞれ10個のF3種子を用いてF3世代を育てた。性質3から7は、性質1、2とは異なり、F3植物体を育てて初めて評価が可能である。メンデルは、100のF2個体から得たそれぞれ10個体のF3世代を育てて、F2が安定親型であったか雑種型であったかを調査している。雑種型個体と安定親型個体の出現数を見てみると、それぞれで64: 36、71: 29、60:40、67:33、72:28であった。供試数が少なく、性質6以外では2:1の分離比からのずれが見られ、特に性質5では60:40とずれが大きかった。そこで性質5について再度実験を繰り返し、今度は65:35(1.9:1)という分離比を得た。したがって、平均の分離比は確かに2:1で一定であった。実験結果全体をまとめて見ると、F2で顕性を示した個体の3分の2が雑種型で、残りの3分の1はすべて安定親型の顕性を示し、その比は2:1の近似であった。
メンデルは、ここで、F2で観察された顕性と潜性の割合3:1は、正確には雑種型2 : 顕性の安定親型1 : 潜性の安定親型1の割合であると説明する。すなわち、雑種当代F1の自家受粉で得たF2世代の子孫の性質を雑種型と安定親型に別けてみると、それらの比は2:2であり、安定親型では顕性と潜性が1:1の割合で生じ、全体として2:1:1となると説明する。雑種F1の中で合一した両親の性質が、F2ではそのまま変化することなく安定親型として雑種型と同数だけ出現するという事実は、メンデルの粒子説の根幹をなす法則である。これは現在、メンデルの分離の法則(第一法則)と呼ばれている。
7章)雑種のその後の世代
雑種の子孫がF2およびF3世代で安定親型と雑種型に分かれて展開する割合は、後の世代でも同様であろうと考えたメンデルは、さらに実験を継続し、実験1と実験2ではF7世代まで、実験3と7では、少ない供試数ではあったがF6世代まで、実験4、5と6ではF5世代まで交配を進めて、同様の結果、すなわち、雑種型:顕性安定親型:潜性安定親型が2:1:1の割合であったと結論している。
ここで、メンデルは遺伝学を学ぶ学生諸君にはおなじみの数式を導入する。ある1対の対照的な性質について、二つの安定親型のうちで顕性の性質をAで、潜性の性質をaで表せば、両者が合一した雑種の性質はAaで、この二つの対立的な性質が関与する交配子孫の系列は2Aa + 1A + 1aという展開級数で現されると説明する。なお、ここで注意が必要である。メンデルは表現型と遺伝子型を同一記号で表し、雑種型を2つの記号で、安定親型を一つの記号で表している。現在の遺伝学の教科書では、対立遺伝子を顕性のAおよび潜性のaとイタリックで表記し、従って雑種F1の遺伝子型はAa、F1が作る雌雄の配偶子の遺伝子型はAとaだから、F2における遺伝子型は、(A + a)x (A + a)の展開項として1AA + 2Aa + 1aaと表され、表現型は3[A]:1[a]と表されることになる。メンデルの表記法2Aa + 1A + 1aは、1種類の雑種型と2種類の安定親型が出現することを強調したものであろう。
なおメンデルは、雑種が作る子孫の半数は雑種型で、残りの半数からは両親のそれぞれの性質を備えた安定型が1:1の比で出現するという結果に基づいて、自家受粉を続けて得られる雑種n世代の子孫では、全体としてA:Aa : aが2n-1: 2 : 2n-1となることを導きだしている。現在の世代の表記法に従ってメンデルの式を書き改めれば、AA:Aa : aaの比は2(n-1) -1 : 2 : 2(n-1)-1となる。従って、メンデルの雑種第10世代(現在の表記法でF11世代)では、2,048個体のうち1,023個体がそれぞれ顕性か潜性の安定親型で、雑種個体はわずか2個体となると予想できる。この予想は、自家受粉を続ければ、子孫は安定な両親型に回帰し、雑種型は急速に失われるとしたケールロイターやゲルトナーたちの予想と結果的に同じだが、メンデルはこれに確固とした実験的なデータの裏付けを与えたことになる。これは、19世紀の終わりにデンマークのウィルヘルム・ヨハンセンが提唱した「純系説」の先取りであった。ヨハンセンは自殖性のインゲンを用いて、重い豆と軽い豆を選抜し、それぞれ自家受粉を繰り返して得た重い豆あるいは軽い豆をつける純系集団を調査し、それぞれの純系内では種子の重さは正規分布を示すことを明らかにした。ヨハンセンの唱えた「純系説」は、ダーウィンの主張した選抜は、純系内では効果がないことを明らかにしている。すなわち、多様性をもつ雑種こそが選択の可能性の基礎であるという事実は、多様性がなければ選択はありえないという意味で進化を考える際の重要な原則である。
8章)複数の性質に関する雑種の後代
ここまでは一つの対照的な性質に着目した実験だったが、メンデルはさらに進んで、対照的に異なる複数の性質を併せ持つ両親間の雑種でも同じような伝達様式が見られるかどうかを知ろうと試みた。この場合には、雑種の性質は、より多くの顕性の性質をもつ一方の親型にいつでもほぼ完全に同一となることが実験の結果から明らかだった。例えば、丈が低く、頂性で白い花をつけ、膨らんだ莢をつける個体を種子親にして、背が高く、腋性で紫の花をつけ、くびれた莢をつける個体を花粉親にした場合には、雑種は顕性の膨らんだ莢をつける点でのみ種子親に似ることになる。もし一方の親が顕性の性質のみを示す場合には、雑種はその顕性親とほとんど区別できないか全く同じ性質を示すことになる。
実験1)種子の形と子葉の色に関する2因子交配ついて
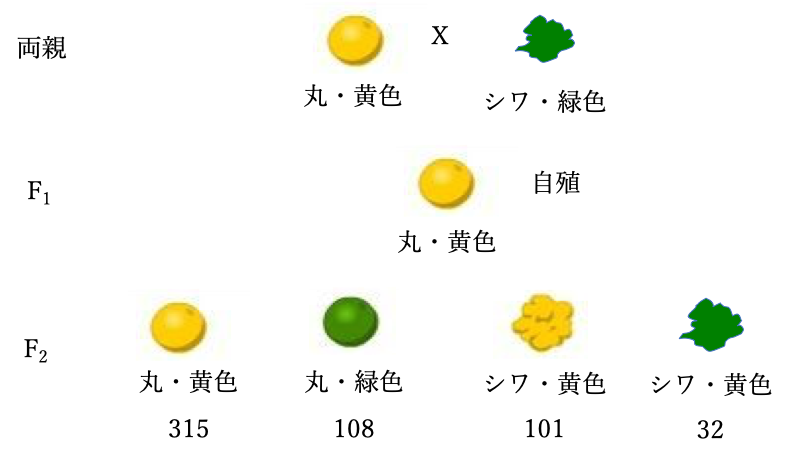
丸・黄色種子を着ける母親(種子親)と、シワ・緑色種子を着ける花粉親の交配で得たF1の丸・黄色種子から生じた15個体を自殖して、556個のF2世代の分離を調べたところ、丸・黄色種子が315個、丸・緑色種子が108個、シワ・黄色種子が101個、シワ・緑色種子が32個得られた(図4)。
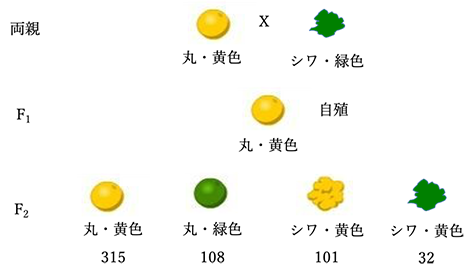 図4 二因子交配におけるF2世代の分離
図4 二因子交配におけるF2世代の分離
|
両親の性質の組み合わせからなる4種類の性質を示したF2種子の全てからF2世代の植物体を育て、さらにF2植物の自家受粉で得られたF3世代の種子についてそれぞれ調べた。植物体を得ることができなかった11個の丸・黄色種子と、着粒しなかった植物体3個体を除いた結果は以下の通りだった。すなわち、二つの性質についてともに顕性の丸・黄色F2種子からは301個体のF3世代が得られ、そのうち38個体は丸・黄色種子のみを着粒したが、65個体からは丸・黄色と丸・緑色が、60個体からは丸・黄色とシワ・黄色が、138個体からは丸・黄色、丸・緑色、シワ・黄色とシワ・緑色の種子が分離した。さらに、シワ・黄色種子から得た96個体のF3世代のうち28個体はシワ・黄色種子のみをつけ、68個体からはシワ・黄色とシワ・緑色の種子が得られた。同様に、108個の丸・緑色種子から得た102個体のF3世代のうち35個体は丸・緑色種子のみをつけ、67個体からは丸・緑色種子とシワ・緑色種子が得られた。シワ・緑色種子から得た30個体がつけた種子は全てシワ・緑色種子で安定親型と同一であった。
ここで母親のもつ二つの顕性の性質をそれぞれA, Bで表し、父親の二つの潜性の性質をa, b、雑種の性質をそれぞれAa, Bbで表せば、上で得られたF3の結果は、二つの性質がともに顕性のA, Bを示した計301個体では、そのうち38個体がAB、65個体がABb、60個体がAaB、138個体がAaBbであったことになる。メンデルは残りの3種類、すなわちどちらか一方の性質のみ顕性のグループ(Ab, aB, aBb, Aab)に属する198個体と両方の性質について潜性のグループ(ab)30個体も含めて同様の解析データを示した上で、全体を9種類のグループに分けた。その内訳は以下の通りである。
38個体がAB、35個体がAb、28個体がaB、30個体がab、65個体がABb、
68個体がaBb、60個体がAaB、67個体がAab、138個体がAaBb
さらに、この結果を安定親型と雑種型で分類したメンデルは、全体を以下の3つの基本的なグループに分類している。
第1群 AB, Ab, aB, ab (両方の性質が安定親型) 平均33個体
第2群 ABb, aBb, AaB, Aab(一方が安定親型、他は雑種型) 平均65個体
第3群 AaBb(両方が雑種型) 138個体
第1群を構成する4種類のうちABとabは両方の性質について顕性の安定親型か潜性の安定親型であり、AbとaBは一方について顕性の安定親型で他方は潜性の安定親型であって、各種類がそれぞれ平均して33個体ずつ現れたことになる。第2群を構成する4種類ABb、aBb、AaBとAabはどれも一方の性質について顕性または潜性の安定親型で、他方については雑種型であって、4種類がそれぞれ平均65個体ずつ現れた。最後の第3群は両方の性質について雑種型で、その出現数は138個体であった。
従って、3つの群を構成する個体数の平均比率は1 : 2 : 4の近似であったことになる。さらに、以上の3群を構成する9種類の各項をまとめると、(AB + Ab + aB + ab)+(2ABb + 2aBb + 2AaB + 2Aab)+(4AaBb)となることから、メンデルは、二つの性質の組み合わせシリーズの各項は(A + 2Aa + a)x(B + 2Bb + b)の展開項であると結論する。
メンデルは論文中で記述していないが、これを表現型に従ってまとめれば、二因子交配におけるF2世代の分離比は、おなじみの[AB] :[Ab] : [aB] : [ab] =9 : 3 : 3 : 1となる。
実験2)実験1の2種類の性質(種子の形と子葉の色)に、第3の性質である花の色(すみれ色の花か白色の花)を加えた3因子交配について
解析の対象となる因子の数が3つだから当然だが、メンデルは、全ての実験のうち、この実験に最も多くの時間と労力をかけた。実験1)と全く同じ解析と推論から、3種類の異なる性質を合わせもつ雑種24個体に着粒した687個の種子と、それらを育てて得た639の植物個体を対象に、交配で得た全体の結果を以下の様にとりまとめた。
すなわち、花の色に関する顕性の性質をCで、潜性の性質をcで表すと、全体のシリーズは、(A + 2Aa + a)x(B + 2Bb + b)x (C + 2Cc + c)の展開項として、次のようになる。
ABC + ABc + AbC + Abc + aBC + aBc + abC + abc +
2ABCc + 2AbCc + 2aBCc + 2abCc + 2ABbC + 2ABbc +
2aBbC + 2aBbc+ 2AaBC + 2AaBc + 2AabC + 2Aabc +
4ABbCc + 4aBbCc + 4AaBCc + 4AabCc + 4AaBbC + 4AaBbc +
8AaBbCc
なお、これを表現型に従ってまとめれば、 [ABC] : [ABc] : [AbC] : [aBC] : [Abc] :
[aBc] : [abC] : [abc] = 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1となる。
メンデルは、実験2)でも実験1)と同様に、展開数列の27項を含む上記の結果を安定親型と雑種型に着目して分類し、両親が3対の性質・形態について異なる雑種子孫の展開は全体として以下の4つの群に分類されるとした。
・3つの性質がすべて安定親型:(ABC, ABc, AbC, Abc, aBC, aBc, abC, abc)
・2つが安定親型で1つが雑種型:(ABCc, AbCc, aBCc, abCc, ABbC, ABbc,
aBbC, aBbc, AaBC, AaBc, AabC. Aabc)
・1つが安定親型で2つが雑種型:(ABbCc, aBbCc, AaBCc, AabCc, AaBbC, AaBbc)
・3つ全てが雑種型: (AaBbCc)
各群の平均構成数はそれぞれ、10個体(7から14)、19個体(14から25)、43個体(36から49)、78個体で、これは10個体:20個体:40個体:80個体の近似であり、比率は1 : 2 : 4 : 8に近い。
メンデルは他の形質についても、供試数は少なかったが、2つおよび3つの因子を組み合せた実験を行い、ほぼ同様の結果を得たと報告している。メンデルは、「複数の明確に区別できる性質を組み合せた雑種の子孫では、対となる性質の一連の組み合わせ項が現れるが、それらは両親がもつそれぞれ対をなす異なる性質の互いに独立な組み合せの結果として生じる」とする結論を導いている。これは現在、私達がメンデルの独立の法則(第二法則)と呼ぶものである。
メンデルは、以上の推論から、両親で異なる性質の数がn対あるとすれば、雑種で生じる組み合わせシリーズの構成数は3n、シリーズに属する個体の数は4nで、そのうち安定親型は2nであると予想する。
その上で、7対の異なる性質を対象にしたエンドウでは、独立の法則に従って期待される27 = 128種類の安定親型の全てが実際に得られたと述べ、この結果は、「ある植物種の複数系統で見られる安定な性質が、人為的な自家受精を繰り返すことで、法則に従って可能な全ての組み合わせとして取得可能である」ことの具体的な根拠であると結論している。
9章)雑種が作る生殖細胞
メンデルはさらに、雑種の子孫に現れる顕性あるいは潜性の安定親型および雑種型と雑種が作る配偶子(卵細胞と花粉)との関係を明らかにする目的で、それまでに得られたすべての結果に考察を加え、重要な仮説を提唱する。この仮説は、「雑種の子孫では、安定親型が全ての可能な組み合わせで出現する」という観察結果に基づいて提唱されている。メンデルの推論と導かれた仮説は以下の通りである。
「雑種の子孫では可能な全ての組み合わせをもつ安定な親型が現れる。安定な親型は胚珠で作られる卵細胞と葯で作られる花粉細胞が同じ性質をもつ場合にのみ可能であるから、雑種の子孫で(様々な組み合わせを持つ)安定な親型が生じるためには、どちらの配偶子でも、正確に同じ性質を備えた因子の関与がなければならない」。この記述から、私達は、安定な親型の性質を示す子孫(これらを様々に組み合わせた子孫)が雑種から形成されるためには、当該の性質を決める同種の因子が両親から同時に供給される必要があることをメンデルが正確に理解していたことを知る。さらにメンデルは、「各性質を担う卵細胞と花粉細胞が雑種の胚珠と葯で同数だけ作られ、それらが無作為に受精する」と推論し、これを確認するために以下の実験を実施した。
メンデルが実施した実験は、種子の形と色に関する以下の4通りだった。ここで、種子親の性質をAB(Aは丸種子、Bは黄色種子)、花粉親の性質をab(aはシワ種子、bは緑色種子)とすれば、各実験は以下のようにまとめられる。
実験1と2は、それぞれ雑種植物体が作るAB, Ab, aB, abの卵細胞を、元の親植物体が作る花粉AB(実験1)あるいはab(実験2)で受精させる交配で、その目的は、雑種が作る卵細胞の構成を明らかにすることであった。
実験3と4は、元の親植物体が作る卵細胞AB(実験3)あるいはab(実験4)を、雑種植物が作る花粉AB, Ab, aB, abで受精させる交配で、その目的は、雑種が作る花粉細胞の構成を明らかにすることであった。
上記の交配様式は、雑種個体と元の親個体をそれぞれ種子親あるいは花粉親として入れ替えて用いた正逆交配である。正逆交配は、雌雄配偶子の効果を評価し、メンデルは認識していなかったが、母性効果(細胞質遺伝の効果)を見出す手段ともなる。一方、戻し交配とは、雑種個体をどちらか一方の親個体で交配する様式を云う。遺伝解析の目的は、純系(ホモ接合体)間の交配で生じる雑種(ヘテロ接合体)の作る配偶子の遺伝子型割合を知ることであるから、特に潜性安定親型(潜性の純系個体)と雑種型との戻し交配が重要である。この交配様式では、次代の子孫の表現型の分離が雑種植物体の作る配偶子の遺伝子型分離を直接に表すことになるから遺伝解析では重用され、一般に検定(テスト)交配と呼ばれている。
以上の4通りの交配実験から、メンデルは以下の結果が得られると期待した。すなわち、雑種植物体が作る卵細胞と花粉の種類がどれも同数だけ作られ、それらの無作為な受精で生じる全ての子孫が生存可能であるとの仮定がなりたてば、それぞれ以下の個体が同数ずつ得られるはずである。
実験1 AB, ABb, AaB, AaBb(すべて丸・黄色種子)
実験2 AaBb, Aab, aBb, ab(丸・黄色、丸・緑色、シワ・黄色、シワ・緑色)
実験3 AB, ABb, AaB, AaBb(すべて丸・黄色種子)
実験4 AaBb, Aab, aBb, ab(丸・黄色、丸・緑色、シワ・黄色、シワ・緑色)
実験1と3の結果および2と4の結果はそれぞれ同一の結果を与えると期待されたが、得られた結果はまさにその通りであった。すなわち
実験1 98個の丸・黄色種子
実験3 94個の丸・黄色種子
実験2 31個の丸・黄色種子、26個の丸・緑色種子、22個のシワ・黄色種子、
26個のシワ・緑色種子
実験4 24個の丸・黄色種子、25個の丸・緑色種子、22個のシワ・黄色種子、
27個のシワ・緑色種子
メンデルは以下のデータを示して、さらに念押しをしている。
実験1で得た98個体のうち90個体、実験3で得た94個体のうち87個体、実験2で得た105個体のうち110個体、実験4で得た98個体すべてを用いて、それぞれの自殖次世代を調べ、以下の結果を得た。
実験1 20個体が丸・黄色(AB)のみ、23個体が丸・黄色あるいは丸・緑色(ABb)、25個体が 丸・黄色あるいはシワ・黄色(AaB)、22個体が 丸・黄色、シワ・黄色、丸・緑色あるいはシワ・緑色(AaBb)の種子
実験3 25個体が丸・黄色(AB)のみ、19個体が丸・黄色あるいは丸・緑(ABb)、22個体が丸・黄色あるいはシワ・黄色(AaB)、21個体が 丸・黄色、
シワ・黄色、丸・緑色あるいはシワ・緑色(AaBb)の種子
実験2 31個体が丸・黄色、シワ・黄色、丸・緑色あるいはシワ・緑色(AaBb)、26個体が丸・緑色あるいはシワ・緑色(Aab)、27個体がシワ・黄色あるいはシワ・緑(aBb)、26個体がシワ・緑色のみ(ab)の種子
実験4 24個体が丸・黄色、シワ・黄色、丸・緑色あるいはシワ・緑色(AaBb)、25個体が丸・緑色あるいはシワ・緑色(Aab)、22個体がシワ・黄色あるいはシワ・緑(aBb)、27個体がシワ・緑色のみ(ab)の種子
まとめれば、以上4種類の子孫の割合は全て検定交配で期待される割合1:1:1:1の近似であった。
メンデルの検証実験はさらに続き、花の色(すみれ色と白色)と茎の長さ(長い茎と短い茎)の組み合わせ、さらに莢の形(膨らんだ莢とくびれた莢)、莢の色(緑色の莢と黄色の莢)と花のつき方(腋性と頂性)の組み合わせについても同様の実験を行っている。2つの性質を対象とした実験では、すみれ色の花・長い茎:白色の花・長い茎:すみれ色の花・短い茎:白色の花・短い茎が47: 40: 38: 41の割合で現れ、4つの性質それぞれに着目すれば、すみれ色の花:白色の花:長い茎:短い茎の割合は85: 81: 87: 79で1:1:1:1の近似であった。3対の異なる性質を対象とした小規模な実験についても、データは示されていないが、期待される結果と完全に一致する結果を得たとメンデルは述べている。
以上の実験からメンデルの下した結論は以下のとおりであった。「エンドウの雑種では、卵細胞と花粉細胞の受精による無作為な結合で可能となる全ての異なる性質の組み合わせが正確にほぼ同数だけ現れる」。
メンデルは、以上全ての実験結果をとりまとめて、以下のような公式化を提案する。メンデルは推論の過程を丁寧かつ厳密に組み立てて繰り返し語っているが、以下では最も重要な点のみを抜粋する。
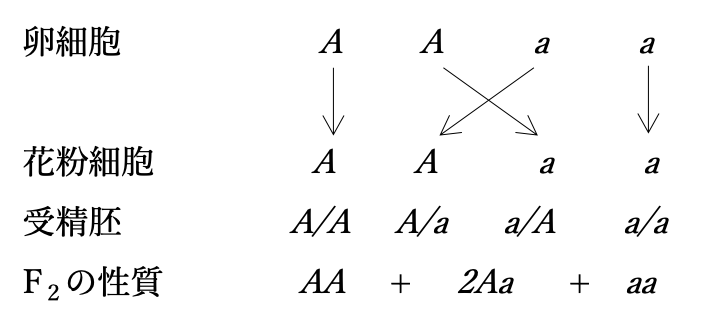
対照的な性質あるいは形態であるAあるいはaをもつ同数の花粉細胞が同様に同数のAあるいはaの卵細胞と無作為に組合わさることで、A/A + A/a + a/A + a/a(分子と分母はそれぞれ花粉細胞と卵細胞の型を表す)の受精胚が確率論的に同数生じる(図5)。
1番目(A/A)と4番目(a/a)は花粉細胞と卵細胞が同一型で、それらの受精で生じた個体は安定親型であり、2番目(A/a)と3番目(a/A)は異なる型の卵細胞と花粉細胞が受精することで生じた雑種型である。卵細胞あるいは花粉細胞がどちらの型であろうとも、A/aとa/Aは雑種型で、表れる性質は同じだから(正逆交雑でも結果は同じであるから)、まとめるとA + 2Aa + aとなる。(図5)に示した結果は、1因子に関する雑種F1の自家受粉によるF2世代の一般化である。もちろん、2種類の卵細胞と2種類の花粉細胞の合一は無作為な偶然の結果である。
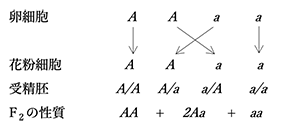 図5 一因子交配で雌雄配偶子の受精から
図5 一因子交配で雌雄配偶子の受精から
生じるF2世代の分離
2世代の分離
|
全く同様の推論から、2因子に関する交配で得られる雑種の子孫は(A + 2Aa + a)x(B + 2Bb + b)の展開項で与えられる。すなわち、雑種の子孫は、安定親型に対応した4種類の同数の卵細胞と花粉細胞、すなわちAB, Ab, aB, abの組み合わせから生じる9種類の異なる組み合わせシリーズである(AB + Ab + aB + ab )+(2ABb + 2aBb + 2AaB + 2Aab)+ (4AaBb)となる。
3つの異なる性質に関する交配で得られる子孫も全く同様で、雑種で合一した対照的な性質は全ての可能な組み合わせから生じる安定親型に対応した8種類の同数の卵細胞と花粉細胞、すなわちABC, ABc, AbC, Abc, aBC, aBc, abC, abcの組み合わせから生じる27種類の異なる組み合わせシリーズの各項、ABC + ABc + AbC + Abc + aBC + aBc + abC + abc + 2ABCc + 2AbCc + 2aBCc + 2abCc + 2ABbC + 2ABbc + 2aBbC + 2aBbc + 2AaBC + 2AaBc + 2AabC + 2Aabc + 4ABbCc + 4aBbCc + 4AaBCc + 4AabCc + 4AaBbC + 4AaBbc + 8AaBbCcで構成されることになる。これは、(A + 2Aa + a)x(B + 2Bb + b)x (C + 2Cc + c)の展開項である。
メンデルの論文では、以上の序論、材料と方法、結果を記述した9つの章に続く以下2つの章で、他の植物種で得た、あるいは他の研究者たちが報告した結果を勘案し、エンドウで得た仮説の一般化の可能性と、種の形成における雑種の意味がそれぞれ論じられている。
10章)他の植物雑種を用いた実験
この章では、他の植物種を用いて実施した雑種実験の結果を説明している。メンデルはまず、エンドウと同様の結果が他の植物種でも得られるかどうかを検証するために、インゲン属(Phaseolus)を用いた実験を開始し、論文執筆時までに完了した2つの組み合わせ、すなわちPh. vulgaris x Ph. nanusとPh. nanus x Ph. multiflorusからなる種間雑種を対象にした3つの対照形質に関する小規模な実験について解説している。近縁の種間雑種Ph. vulgaris x Ph. nanusで得た結果は、エンドウのそれと全く同様であった。主軸が長く(顕性)、成熟するとくびれる黄色の莢をつける(潜性)Ph. vulgaris の性質と、主軸が短く(潜性)、緑色の膨らんだ莢をつける(顕性)Ph. nanus の性質は、子孫でエンドウの実験結果から期待された通りに振る舞い、8種類の安定親型が生じた。一方、形態の大きく異なる種間の組み合わせPh. nanus x Ph. multiflorus では、一様でない複雑な結果が得られた。種子親に用いたPh. nanusは、常に短い主軸の先の総状花序に白い花をつけ、まっすぐで膨れた滑らかな莢に小さな白い種子をつける。対照的に花粉親に用いたPh. multiflorusは、丈が高くからみつく茎、極めて長い総状花序に紫がかった赤い花、ざらざらした鎌型に曲がった莢、赤みの強い桃色に黒い斑点やシミが入った大きな種子をつける。花粉親によく似た雑種は、しかしながら稔性が甚だ低く、49粒の種子から44個体の次代しか得ることができなかった。雑種で隠れていたPh. nanus の潜性の性質は、種々の組み合わせで再び出現したが、それらの割合は大きく変動した。それでも、主軸や莢の性質については、エンドウと同様にほとんど3:1に近い分離の結果が得られた。
インゲン属植物の種間雑種の実験を説明したこの章では、特に雑種の稔性が不安定で低いことなどもあって、メンデルは詳細なデータを記載せず、「インゲンの形態について雑種で見られた展開は、エンドウの場合とほぼ同じ法則に従っていた」とする結論だけを述べている。しかし、ここでメンデルは以下の点に気づき記録している。
インゲンの花の色は紫色から薄紫色、白色まで変化する。メンデルは、白色と紫色を交配した雑種第二代では、開花した31個体のうち白色の花をつけた個体が1個体しかなかった事実を述べ、期待される4分の1から大きくはずれたこの比の出現を次のように説明した。花の色という性質が全体として紫色がかった赤色を生じさせる独立なふたつの(顕性)因子A1とA2で決まっているとするならば、(潜性)因子の白色aとの交配からそのような低い頻度で白色が出現することが説明できる。例えば、二つの因子A1とA2のどちらか一つを持ちさえすれば赤紫色となると仮定すれば、1A1A2 +2A1aA2 +1A2a + 2A1A2a + 4A1aA2a + 2A2aa + 1A1a + 2A1aa + 1aaの構成から、16に1の割合で白色の花をつける個体aaが得られることになる。ただし、二つの赤紫色を決める因子A1とA2か関与するとすれば、白色因子aにもa1とa2の二つがあるはずで、上の展開級数は1A1A2 +2A1a1A2 +1A2a1 + 2A1A2a2 + 4A1a1A2a2+ 2A2a2a1 + 1A1a2 + 2A1a1a2 + 1a1a2となるだろう。メンデルの見た分離比は現在の知識によれば、後にウィリアム・ベイトソンがエピスタシスと名付けた遺伝現象、すなわち、複数の遺伝子座にある複数の遺伝子間の相互作用が同一形質に関与する現象として説明できる(Bateson, W. (1909) Mendel’s Principles of Heredity. Cambridge University Press, London, UK.)。エピスタシスの本来の(古典遺伝学的)意味は以下の通りである。それぞれ固有の表現型を支配する遺伝子Aの変異型とBの変異型を同時に持つ個体が変異型の表現型Aを表す時、遺伝子AはBの上位(エピスタティック)、遺伝子BはA の下位(ハイポスタティック)の関係にあることを云う。現在では、より広い意味で、ゲノム中の一つの多型がある形質に及ぼす効果が他の多型に依存して決まる遺伝現象を云う(第4章参照)。メンデルはインゲンで、花の色という形質が2つの遺伝子(同義遺伝子)の関与で決まり、通常の分離比の9:3:3:1ではなく15:1が観察される重複遺伝子作用の例をみたのであろう。メンデルは結果を正しく解釈していたと思われるが、論文中では、「ここで試みた説明には、実験上の支えは何もなく、単なる推定に基づくものであることを忘れてはならない」と注意深く述べている。
続いて、メンデルは、「多くの観賞用園芸植物の花色は極めて変異性に富んでいるが、確かなことはまだ何も分かっていない」と述べるが、観賞用の園芸植物について育種家の間で一般に言われていた「種の安定性は栽培によって高度に撹乱されるのであって、栽培型の展開は規則性のない偶然性の高いものである」とする主張に強く反論している。常に抑制的なメンデルも、この点では極めて辛辣である。メンデルは、一方で「生育条件が変えられ、さらにその種が新しい環境に適応する能力を持つならば、性質・形態の変化はどこでも起こるはずである」、「栽培が新しい種の形成に好都合であって、自然条件下では失われてしまうはずの多くの変種(品種)が人の手によって得られるであろうことを、認めないわけにはいかない」として育種家の主張を部分的ではあるが認めた上で、「庭園の土壌へ植物体を移植するだけで、何故そのように決定的で持続的な変化がもたらされるのか全く不明である」、「自然に開かれた土地における植物の展開が、庭園の苗床におけるそれとは別の法則によって支配されていると真面目に主張する者はあるまい」、ましてや「変種・品種を形成する傾向が、その種が速やかにあらゆる安定性を失い、その子孫が高度に異なった形態からなる無限のシリーズへと分かれていくほどに大きく増加するという仮定を正当化する証拠は一つもない」と主張する。メンデルは、変種(品種)は種と同様に一定の安定性を備えた存在であると考えている。実際、メンデルは論文の至るところで、合計96回も、「安定な性質」、「一定な子孫」、「一定な組み合わせ」、「一定な現れ」「一定な法則」、「一定な種」など安定あるいは一定という述語を多用している。
さらにメンデルは以下のような議論を展開している。「栽培条件の変化が変異の唯一の原因であるとするならば、何百年も同一環境下で栽培されてきた園芸植物は全て安定性をとり戻しているはずであるが、実際にはそうではなく、そうした環境でも多くの変種や可変性のものが見られる」、「自殖性のエンドウ属やインゲン属のようなマメ科植物は例外的ではあるが、そうした植物種でも多様な環境下で千年以上も栽培されてきた間に多くの変種が生まれている。しかもこれらの変種は、変化のさして著しくない栽培環境の下でも、野生環境で生育する種と同等の安定性を維持している」。メンデルのこうした主張は、ダーウィンの進化論との関係で重要であるから、本書の第5章と6章で改めて考察する。
メンデルは、「栽培植物の変異性については、今までほとんど注意が向けられてこなかった一つの要因が働いていると言ってまず間違いがない。様々な実験結果から、栽培植物はほとんど例外なく、種々の雑種展開数列の各項であって、頻繁な交配自体によって変化しその発達が法則に準拠して変化したり中断したりすると結論できる」、また「栽培植物は、多くの場合、多数が近接して植えられるから、変種間、品種間あるいは種間で自然に起こる相互交雑にとって好都合な条件が与えられていることに注意しなければならない」と指摘している。メンデルの控えめな主張はおそらく、栽培植物の多様性に預かる雑種の重要性を軽視してはならないと云う主張であったのであろう。
11章)結語
論文を締め括る最終章では、メンデルがエンドウで得た結果を、ドイツの植物学者ジョセフ・ゴットリープ・ケールロイターと植物学者・医師カール・ゲルトナーがタバコ属2種の種間雑種Nicotiana paniculata x Nicotiana rustica で得た結果と比べたうえで、雑種と種の形成の関係について論じている。メンデルは、二人の先人が得た結果を次のようにまとめている。「雑種の外観形態は、両親の中間型か、あるいは、どちらか一方の親により似ているか、またはそれとほとんど見分けがつかない場合もある。通常、一回の自家受粉で得た後代の大多数は雑種型を維持するが、種子親あるいは花粉親に近いものもわずかながら存在する。しかし、すべての雑種に例外なくこれが当てはまるわけではない。子孫の一部あるいは全体が両親の一方あるいは他方に近づく場合があり、他方で完全な雑種型として安定に維持されるものも見出せる。変種間の雑種にも種間雑種と同様の振る舞いが見られるが、変種間雑種ではより著しい形態の多様性とともに親型に戻るより甚だしい傾向が認められる」。
以上のタバコの結果は概ねエンドウの結果と一致しているとメンデルは結論する。しかし彼らが、「雑種とその後代では様々な傾向が認められたというだけの不明確な観察結果の記述にとどまった理由の一つは、ゲルトナー自身が認めていたように、解析の対象とした性質や形態の評価が多くの場合に困難であったことに帰せるだろう」と推察している。さらに、「親型と雑種型の現れ方に明確な規則性が存在することに二人が気づかず、明快な結論を下せなかった大きな理由は、多数の性質について互いに異なる親植物を交配に用いたこと、一方で解析した子孫の数が十分大きくなかったことに原因がある」と推定する。エンドウの結果に従えば、「個々の実験で生じる展開数列の項の数は3の累乗で増加するから、性質や形態の数が多くなるほど大きくなるはずである。従って、調査する子孫の数が少なければ正確な評価はできない」。両親間で相違する性質がn種類あれば展開数列の項の数は3nで、種類に属する個体の数は4nとなって急激に増加する。従って、「例えば、7つの性質・形態のすべてに相違のある両親から得た雑種を考えた場合、展開数列の項は2,187種類で16,834個体を含むはずであり、結果は観察者が選んだ偶然に大きく作用されるだろう」。この場合には、「16,000個体以上の雑種子孫のうち両親型はそれぞれ1個体現れるに過ぎない」。実際、ゲルトナーによれば、雑種の子孫では両親型そのものではなく、それに比較的類似した個体が得られたに過ぎなかった。
メンデルは続いて、ゲルトナーがオダマキ属(Aquilegia)、ハナアオイ属(Lavatera)、ダイコンソウ属(Geum)およびナデシコ属(Dianthus)などの雑種で、ヴィフューラがヤナギ科の雑種で、特に優れた稔性を示す組み合わせの場合に観察した結果の重要性について、そこでは本質的に異なる特別な状況が見られると指摘する。すなわち、こうした雑種では安定で純粋種のように繁殖する子孫が存在するとしたゲルトナーの観察を引用して、「この類の安定な雑種は新たな種としての存在意義を獲得するという意味で、植物の進化の歴史にとって極めて重要である。この事実の真実性は、最も優れた観察者たちによって正当化されており、疑うことはできない」と述べている。事実、ゲルトナーは、庭園で自殖繁殖するナデシコ属の雑種を10世代まで観察してこのことを報告していた。
続くページでメンデルは、エンドウのような雑種の子孫が可変的である場合とゲルトナーが観察した安定な雑種が維持される場合の違いが生じる原因の説明を試みているが、その説明にはかなり苦慮している。メンデルは、「雑種が成立するためには、雌雄の配偶子の間に何らかの調停が行われる必要がある」として、次のように説明する。「エンドウでは、雑種が様々な卵細胞と花粉細胞を形成することが子孫の可変性の理由であるが、雑種が自殖を通じて安定なままであるような例では2種類の生殖細胞がどちらも同一で雑種である母親の基となる基礎細胞あるいは調停細胞と同じ場合が想定されるのである」。メンデルは、受精卵の中で、相違する因子の間の調和を図る何らかの調停が基礎細胞あるいは調停細胞で行われるのだろうと考えている。確かに、こうした仮定が成立すれば、雑種の自殖子孫は雑種の母親植物と同じ性質あるいは形態を示すことになるだろう。しかし、雑種はそもそもヘテロ接合であり、減数分裂を通じてそれらから雌雄で同一遺伝子型の配偶子が生じ受精で結合することは、関与する因子の数が多ければ多いほど、極めて稀にしかありえない。
実は、インディアナ大学のラルフ・クレランドの細胞遺伝学的な研究から1922年になって初めて、ヘテロ接合型の親植物の自殖子孫で常にヘテロ性が持続されるような特殊な現象が存在することがわかった。それは、ユーゴ・ド・フリースが突然変異を発見したオオマツヨイグサで見られた。オオマツヨイグサ(Oenothera lamarkiana)はヘテロ接合であるが、常に純系のように振る舞い同じ子孫を作る。オオマツヨイグサの作る配偶子は発見者のオットー・レナーによってそれぞれゴーデンスとヴェランスと名づけられた2種類の複合体を構成しており、受精によって生じる子孫はゴーデンスとヴェランスの組み合わせのみで、ゴーデンスとゴーデンスおよびヴェランスとヴェランスの組み合わせは生存できない。これには、オオマツヨイグサの14本の染色体のうち12本が互いに転座した相互転座と呼ばれる染色体の大規模な構造変異とそれぞれの複合体に致死遺伝子が存在することで起こる平衡致死と呼ばれる現象が関与している(Steiner, E. (1982) Ralph Eiskine Cleland 1892-1971. National Academy of Science, Washington D.C.; Sturtevant, A.H. (1926) Renner’s studies on the genetics of Oenothera. The Quarterly Review of Biology 1: 283-288.)。メンデルの想定がこの例には当てはまるか否か分からないが、メンデルの考案した調停細胞と云う概念は当たらずとも遠からずと考えられるのかも知れない(種間の雑種が雑種性を保ったまま新たな新種として直接的に生じる場合が知られており、それは非還元配偶子間の融合によって生じる異質倍数性である。その代表例はコムギ属で、例えば、4倍性のマカロニコムギはBBゲノムを持つ2倍性の野生種とAAゲノムを持つ2倍性の栽培種の自然交雑から生じたが、雑種のゲノム構成がABのままであれば稔性のある新種としては成立しえない。こうした雑種は一代限りであることから一代雑種と呼ばれ、ウマとロバの間で生まれたラマ、ライオンとヒョウの間のレオポン、トラとライオンの間のライガーなどが知られている。ところがコムギでは、雌雄の減数分裂で単数性の配偶子ではなくそれぞれ2倍性のAAゲノムとBBゲノムを持つ非還元配偶子が稀にでき、それらが受精で融合してAABBゲノムを持つ稔性のある雑種として新たな異質4倍体のマカロニコムギが生まれた。異質6倍体でAABBDDゲノムを持つパンコムギもAABB種とDD種の間で起こった同様のプロセスを経て生まれ、人間がこれを選んで育てたのである。こうした異質倍数性は、イネ科やアブラナ科など多くの植物で知られている)。
原著論文に戻ろう。メンデルは、ゲルトナーが、上記の4属に加えてタバコ属、センノウ属(Lychnis)、ゼニアオイ属(Malva)とマツヨイグサ属(Oenothera)を対象に実施した「人為的な交配による、ひとつの種の別の種への変換」実験の結果に考察を加えている。ゲルトナーが確認した現象、すなわち「雑種は、一方の親を用いて繰り返し交配を続けると、交配親の性質に戻る」とした現象を、エンドウの結果に基づいて、次のように説明している。「3つの性質について異なる両親間の交雑で、種ABCを種abcへ変換する場合を考える。種ABCを種abcの花粉で受精させた雑種第一代では、ABC, ABc, AbC, aBC, Abc, aBc, abC, abcからなる8種類の卵細胞ができる。この卵細胞をabcの花粉細胞でもう一度受精させると、対応した8種類の子孫、AaBbCc, AaBbc, AabCc, aBbCc, Aabc, aBbc, abCc, abcが生じる。それぞれの項でabcをもつものが一つあるから、どの性質についても、abcによる第2の交配で変換が完了する可能性がある」。実際には、「実験に用いる植物体の数が少なければ少ないほど、両親で異なる性質の数が多ければ多いほど、変換には花粉親abcによる多くの交配が必要となり、ゲルトナーが観察したように、同じ種で、一世代か二世代の遅れが簡単に起こり得るだろう」。続けてメンデルは、潜性の性質を多く持つ一方の親型への変換は容易だが、顕性の性質を多く持つ親型への変換には時間がかかることを、エンドウで得た結果、すなわち、顕性を示す個体は顕性の安定親型と雑種型の2種類からなるという結果に基づき、論理的に説明している。ゲルトナーの記述した現象は、現在ではひとつの種のもつ遺伝子が他種の遺伝子プールに侵入するイントログレッション(introgression, 遺伝子侵入)と呼ばれる現象として知られるが、メンデルは雑種の子孫から新たな種や変種が生じる可能性を見ていたのであろう。なお、現在では、連続戻し交配と呼ばれる方法によって、ある種Aの核の遺伝構成(ゲノム構成)を別種Bのそれで置き換えることが可能である。連続戻し交雑の回数をnとすれば、種Aの核ゲノムが種Bの核ゲノムに置換される割合は(1 - 1/2n)で現される。連続戻し交配によれば、特定の核ゲノムを種々の細胞質ゲノムと組み合わせた細胞質置換雑種の作成が可能である。コムギ属ではこうした人為的な核置換が広範な種間で行われ、種々の実験系統の作出に有効活用されている(第4章の細胞質遺伝を参照)。
メンデルがエンドウの交配実験で得た結論は、人為的な交配による雑種の子孫では、法則(分離の法則と独立の法則)に従って、対照的な個々の性質について、安定な雑種型(固定した組換え型)を含む可能な全ての組み合わせが生じることであった。この結論に基づきメンデルは、トランスフォーメーションを通じて新たな種が不断に生じるとして種の安定性に異議を唱えていた博物学者達に対するゲルトナーの反論に論評を加えている。ゲルトナーは、人為的な交配によってひとつの種が別の種へ完全に変換できるとする実験結果を、むしろ「種はそれを越えては変化することができない固有の限界内に閉じ込められている」とする見解の証拠として捉えていた。メンデルは、「ゲルトナーの見解を無条件に受け入れることはできないとしながらも、他方で、栽培植物の変異性に関して述べられた推測の注目すべき証拠が、ゲルトナーの諸実験の中に見出される」と述べ、次の文章で論文を締め括っている。「ゲルトナーの実験には、オダマキ属(Aquilegia atropurpureaとcanadensis)、ナデシコ属(Dianthus Caryophyllus、chinensisとjaponicus)、タバコ属(Nicotiana rusticaとpaniculata)のような栽培植物の雑種が含まれるが、これらの雑種は全て、4ないし5回の雑種結合の後でも雑種としての独自性を失ってはいなかった」。
| <<前の章 |
TOP |
次の章>> |
|
Copyright (C)
2021 Chiharu Nakamura All Rights Reserved.
|
|