ヨハン・ヴォルフガング・フォン・ゲーテ
「遺伝の法則」を明らかにした当時もその後も、メンデルは決して科学の中心に身を置く研究者ではなかった。カトリック教会・修道院の一司祭であったメンデルを重要な科学的知見の発見に導いた成功の秘訣は一体何だったのだろう。
本章の前半では、メンデルが「植物雑種の実験」で明らかにした新しい概念の枠組みとしての「遺伝の法則」を要約したうえで、メンデルの遺伝法則の意義と普遍性を多くの拡張例を上げて示す。後半では、メンデルを創造的な発見に導いた秘訣は、鋭い洞察力に裏打ちされた新しい視点はもちろんのことだが、明確な意図と計画のもとに続けた弛まない努力にこそあったことを確認しようと思う。
メンデルが発見した遺伝法則
メンデルは遺伝形質が親から子に伝達する仕組みを説明する明確で単純、簡潔な法則を初めて明らかにしたが、それは完璧なまでの緻密な計画のもとで実施した「植物雑種の実験」から得られた成果であった。メンデルの動機・意図は、「雑種と種の形成の関係」を知ることにあったが、そのためには両親および雑種当代の性質とその後代への伝達様式を明らかにしなければならないとする確かな目的意識があった。以下では、まずメンデルが発見した3つの遺伝法則を、現在の学術用語を用いてまとめて見る。
1)顕性の法則(Law of Dominance, 第三法則)
生物が表す形質は、雌雄の配偶子に由来する一対の対立遺伝子の組み合わせに従って決定される(一般に対立遺伝子と訳されてきたalleleという術語は現在ではアレルとカタカナで表記される。アレルは、染色体上の特定の遺伝子座に存在する変動あるいは多様性「どちらもvariation」を意味するが、近年では特定座位にあるDNA分子の持つ配列の違いを表す用語として、遺伝子以外にも用いられている。しかし、メンデルの扱った形質はまさに特定の形質に対応した対立形質であり、それを担う因子は対立遺伝子であるから、メンデルを述べる場合には従来のまま対立遺伝子という用語を用いることにする)。一対の異なる対立遺伝子についてどちらもホモ接合(pure-line、純系)の両親間の交配による雑種第一代F1(Fは子孫を意味するfilial)ではどちらか一方の親の形質のみが表れる。F1で表現型として表れる形質を顕性(dominance、優性)、隠れて表面に表れない形質を潜性(recessive、劣性)と呼ぶ(優劣を意味するとの誤解を招きかねないという懸念から優性という用語を避けて考案された顕性の法則(第3章の7参照)は、雑種がどちらか一方の親の性質を均一に表す事実に基づき、これを均一性の法則と呼ぶこともあった。なお、雑種も相応しくない用語であるとして、英語のhybridをそのままカタカナでハイブリッドと表記する研究者も多い)。ここで、顕性対立遺伝子をイタリック体の大文字A、潜性対立遺伝子を小文字a、顕性の表現型を[A]、潜性の表現型を[a]で表せば、[A]は遺伝子型AAのホモ接合型およびAaのヘテロ接合型の2種類からなり、[a]は全て遺伝子型aaのホモ接合型である。
2)分離の法則(Law of Segregation, 第一法則)
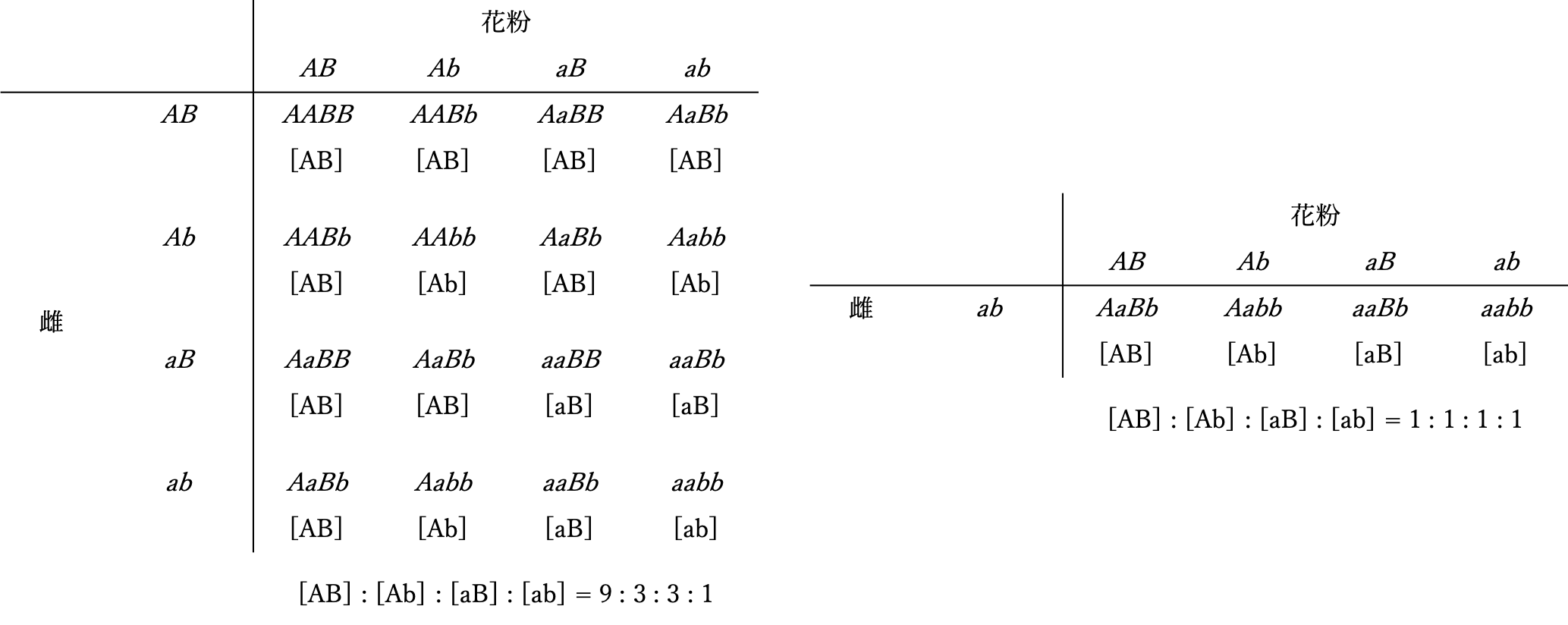
メンデルの遺伝法則のうち最も重要で普遍的な意義を持つ法則である。雑種F1の自殖次代F2では、顕性形質とともにF1で表れなかった潜性形質が一定の割合で再び表れる。すなわち、顕性と潜性の対立遺伝子はF1による雌雄の配偶子形成に際して1:1の比で分離し、続く無作為な受精を通じてF2で再び結合する(自殖または自家受精を行わない、あるいは自殖が不可能な繁殖様式をとる生物種の人為交配では、問題とする対立遺伝子についてともにヘテロ接合である雌雄個体間で交配が行われる。例えば、ショウジョウバエでは、F2はF1のオスとメス(姉弟あるいは兄妹)の交配で得た孫に相当する)。従って、F1が作る雌性配偶子と雄性配偶子の無作為な受精により生じるF2世代の各個体の遺伝子型は(A + a) x (A + a)を展開した各項として与えられ、その結果、AA : Aa : aaが1 : 2 : 1の割合で生じ、表現型では顕性の法則に従って顕性[A]と潜性[a]の割合が3 : 1となる。潜性形質を示す個体は全て遺伝子型aaのホモ接合型であるが、顕性形質を示す個体の遺伝子型はAAのホモ接合型とAaのヘテロ接合型の2種類で、その比は1:2である。なお、F1を潜性ホモ親で戻し交配(検定交配)した世代BC1では、[A] : [a] = Aa : aa = 1 : 1の結果が期待される。問題とする遺伝子についてヘテロ接合であるF1を、aaホモ接合である一方の親と交配する様式を検定交配と呼ぶことは第3章で説明した。顕性の法則に従えば、顕性形質を示すF2個体の遺伝子型を表現型から直接知ることはできないから、これを明らかにするためにはF3世代の解析が必要となる。一方、検定交配によれば、F2における形質の分離比がF1の作る配偶子の遺伝子型の分離比と理論上等しくなることから、ヘテロ接合の個体が作る配偶子の遺伝子型比率を知る目的で実施される遺伝解析では、この交配様式が有用である。(図1)には、2因子交配のF2とBC1における遺伝子型と表現型の分離を示した。
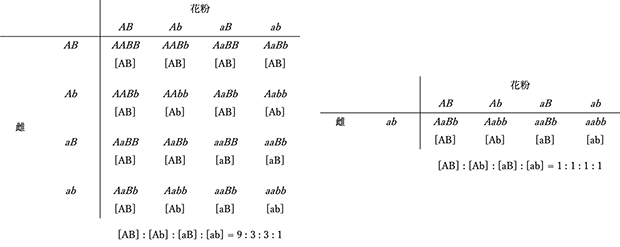 図1 2因子交配におけるF2(左)、BC1(右)の遺伝子型と表現型の分離
図1 2因子交配におけるF2(左)、BC1(右)の遺伝子型と表現型の分離
|
なお、分離の法則から導かれる遺伝的多様性の膨大さをここでみておく。ヒトは男女ともに両親から受け継いだ23対46本の染色体をもち、減数分裂の過程で雌雄配偶子には相同染色体のうち1つが配分される。23対の染色体すべてについて、両親のどちらか一方が1/2の確率で選ばれるから、全体で223(8,388,608)通りの対立遺伝子の組み合わせを持つ配偶子が作られ、それらの無作為な受精によって7.037 x 1013という膨大な数の異なる遺伝子型を持つ子が生まれる可能性がある。換言すれば、私たち一人一人は両親から1/7.037 x 1013 (約70兆分の1)という確率で生まれた。地球上に生きる私たち一人一人は、一卵性双生児を別にすれば皆が異なる遺伝子型を持つ、まさに掛け替えのない存在なのである。なおそのうえ、染色体上の別の座位にある遺伝子は配偶子を作る減数分裂に際して組換えを起こすので、生じる遺伝的な多様性がさらに増大する。メンデルの分離の法則を生み出す減数分裂と受精の遺伝学的意義は明らかであろう。
3)独立分離の法則(Law of Independent Assortment, 第二法則)
2対の対立遺伝子がともにヘテロ接合である雑種F1の自殖次代F2では、各形質に対応した対立遺伝子が独立に分離する。すなわち、ある形質を支配する一対の対立遺伝子は、他の形質を支配する別の一対の対立遺伝子とは独立に分離してF1が作る雌雄の配偶子に入り、それらの無作為な受精を通じてF2に独立に伝達される。従って、2対の対立遺伝子A/aとB/bが関与する2因子交配におけるF2の遺伝子型は、F1が作る配偶子の無作為な組み合わせにより、(AB + Ab + aB + ab) x(AB + Ab + aB + ab)あるいは(AA + 2Aa + aa)x(BB + 2Bb + bb)を展開した各項としてAABB + 2AABb + AAbb + 2AaBB+ 4AaBb + 2Aabb + aaBB + 2aaBb + aabbで与えられ、全体として表現型の分離比は [AB] : [Ab] : [aB] : [ab] = 9 : 3 : 3 : 1となる。この時、それぞれの形質に着目すれば、[A] : [a]も[B] : [b]も依然として 3 : 1の分離比を示す。なお、AaBb x aabbの検定交配では、BC1における表現型の分離比は、AaBbが作る配偶子の遺伝子型の分離比と等しく、AB : Ab :aB : ab と [AB] : [Ab] : [aB] : [ab]がともに1 : 1 : 1 : 1となる。この法則はn対の対立遺伝子についても一般的に成立する。
以上の3つを文章でまとめると、メンデルの遺伝法則は以下のように要約することができる。
2.遺伝子には異なる型をとる対立遺伝子が存在する。
3.遺伝子型は、生物や細胞の遺伝的な構成であり、対を形成する対立遺伝子が互いに異なる遺伝子型をヘテロ接合と呼び、2つの対立遺伝子が同一である遺伝子型をホモ接合と呼ぶ。
4.生物の表す観察可能な特徴を表現型という。ある表現型から、その遺伝子型が直接に分かるとは限らない.顕性の表現型を示す個体の遺伝子型はホモ接合あるいはヘテロ接合である。
5.顕性形質は遺伝子型がヘテロ接合であってもホモ接合であっても表現型に現れるが、潜性形質が現れるのは遺伝子型がホモ接合である場合にのみである。顕性対立遺伝子の存在は潜性対立遺伝子の効果を覆い隠す効果をもつ。
6.対をなす対立遺伝子は生殖細胞(配偶子)の形成時に1:1に分配され、受精時に無作為に組み合わされる。
7.生殖細胞が形成される際には、どの対立遺伝子対も他の対立遺伝子対とは独立に分離する。
メンデルの遺伝法則の拡張
メンデルの遺伝法則、特に分離の法則は両性間の配偶子の受精によって増殖する真核生物で普遍的に成立する遺伝の一般原理である。しかし、顕性・潜性の法則には例外が多く存在する。そうした事象のいくつかについて、代表的な例をそれぞれ挙げて以下で説明する。なお、ここに例として挙げた事象のうちには、見方を変えれば法則の拡張であると考えることが可能な場合も多い。独立の法則については第6章で論じることにする。
共顕性 (codominance):我々の血液型に見られるABO式血液型が代表例であるが、これは集団中で同一遺伝子座に複数の対立遺伝子が存在する場合への拡張例である。メンデルが扱った形質は7つとも、同一遺伝子座に存在する一対の対立遺伝子が支配する対照的な形質だった。しかし現在では、生物集団中には一般に一つの遺伝子座あるいはゲノム中の一つの座位には多数の対立遺伝子(アレル)が存在することが分かっている。ただし、特定の個体が両親から受け継ぐアレルの数は通常2個であって、それらが顕性または潜性の同型であればホモ接合型、顕性及び潜性の異型であればヘテロ接合型と呼んでいる。
ABO式血液型は、メンデルの遺伝法則が再発見された1900年に、オーストリア・ハンガリー帝国ウイーン大学医学部病理学教室のカール・ランドシュタイナーが自分と同僚たちを含めた22人の血液と血清を調べて明らかにした私たちにお馴染みのもので、輸血の出発点になった発見だった( Landsteiner, K. (1961) "On Agglutination of Normal Human Blood". Transfusion. 1 (1): 5-8. ISSN 0041-1132. PMID 13758692; 原著論文はドイツ語、Wiener Klinische Wochenschrift, 46, 1132-1134.)。私たちの血液型は血球表面にある糖タンパク質の糖鎖が抗原となって決められており、血球抗原と反応する血清中の抗体(免疫グロブリン)の間で起こる抗原抗体反応が原因となって血液凝固が起こる。抗原の情報を担う遺伝子は第9番染色体上にあるが、この遺伝子座にはアレルが3つ存在する。血液型A型あるいはB型を決めるA抗原あるいはB抗原はそれぞれのアレルをホモかヘテロの状態でもつ個体で作られる。AB型はA型アレルとB型アレルを同時に持つヘテロ接合であり、O型はどちらの抗原も作らないアレル(ナルアレル)を持つ。一方、O型の血清は抗A抗体とともに抗B抗体を持ち、A型血清は抗B抗体を、B型血清は抗A抗体を、AB型血清はどちらの抗体も持たない。輸血で血液凝固が起こるのは抗原型と抗体型が一致した時のみだから、抗原を持たないO型は万能供血者と呼ばれ誰にも献血可能である。一方、AB型はAB型以外の誰にも献血できないが、誰からも輸血を受けることができる万能受血者である。血液型は関与するアレルの数が3つ以上であり、A抗原を作るアレルとB抗原を作るアレルの間には顕性・潜性の関係が見られない点を除けば、メンデルの遺伝法則が依然として成立している。このようなアレルは共顕性の関係にあると言われる(なお、以下の不完全顕性の項で説明するが、遺伝子型をDNAレベルで判定すれば、全てのアレルは識別可能で共顕性の関係にあると言える)。
不完全顕性 (incomplete dominance):キンギョソウやオシロイバナなどの花色は、遺伝子型がヘテロ接合の個体の表現型が二種類のホモ接合型の中間となる不完全顕性の例である。野生型の赤い花では,一連の酵素反応によってアントシアニン色素が作られる。野生型で活性をもつ顕性アレルによって作られる酵素はホモ接合ではその量がヘテロ接合型より多く,よりたくさんの赤い色素を作るが、不活性な潜性アレルは不活性型の酵素を作り、潜性ホモ接合では花は赤い色素を持たずに象牙色となる。ヘテロ接合では酵素の量がホモ接合に比べて減少し,花中の赤い色素の量も減少するので,希釈効果によってピンク色の花となる。
メンデルが記載した丸種子とシワ種子の違いも実はこうした例の一つである。丸種子とシワ種子の違は、アミロペクチンというデンプンの合成に必要なデンプン枝つけ(または分枝)酵素I(starch branching enzyme I, SBEI)の存在量、したがってSBEIの活性の違いに基づいて決まっている。すなわち、SBEIが十分量だけ存在する場合は種子に丸みのあるデンプン粒が多く含まれ、これが水分を多量に含み、熟すにつれて均一に縮むので丸種子となる。一方、SBEIが全く存在しなければアミロペクチンを欠き、熟すと急速に水分を失い、不均一に縮んで形が不揃いなシワ種子となる。ところで、丸種子の場合は、ホモ型であるかヘテロ型であるかは外観からは判定できない。しかし、顕微鏡観察や生化学的な定量法によって調べてみると、ヘテロ接合体ではデンプン粒の形は中間型であり、アミロペクチン含量も種子がシワにならない程度には高い。外観から評価する形態形質に基づけばホモ接合とヘテロ接合の丸種子は区別できないが、顕微鏡的・生化学的に評価される量的形質に基づけば両者は区別できて共顕性と判定されるのである。さらに、遺伝子の本体であるDNAに基づけば、対立遺伝子の違いはゲル電気泳動法などによって直接に認識することが可能である。メンデルは可視的な外観形質で評価したから顕性を示すホモ接合体とヘテロ接合体を明確に区別できなかったのである。
多面発現効果 (pleiotropy):ある薬剤が期待される効果以外の効果(副作用あるいは副反応)を与える多面効果はよく知られている。本稿を書いている今現在、世界は、新型コロナウイルス(SARS-CoV2)による感染症(COVID-19)蔓延の禍中にあり翻弄されているが、この予防に用いるワクチンにも他のワクチン同様に副反応がある。遺伝子も同様で、単一の遺伝子が複数の遺伝システムあるいは表現型に影響を与える現象を遺伝子の多面発現効果あるいは単に多面発現と云う。メンデルの丸種子とシワ種子は多面発現の一つの例でもあるが、ここではネコの白い毛と青い眼について述べることにする。白い毛と青い眼を持つネコのおよそ4割は難聴である。ネコでは胚発生の間、神経堤に由来する色素細胞は毛包や眼など様々な組織に移動し、毛包ではメラニン色素を合成し、中耳では音を聴くために必須な色素として機能する。これが、白い毛と青い眼の原因となる色素細胞の欠陥が同時に難聴を引き起こす原因となる理由である。
多面発現で最もよく知られているのはフェニルケトン尿症であろう。フェニルケトン尿症(Phenylketonuria, PKU)はアミノ酸のフェニルアラニンを別の芳香族アミノ酸であるチロシンに変換する一つの酵素フェニルアラニンヒドロキシラーゼ(PAH, フェニルアラニン水酸化酵素)の合成を指令する一つの遺伝子の突然変異(mutation)が原因で、それによって酵素の欠損が生じて、体内に過剰なフェニルアラニンとその副産物(代謝産物)が蓄積し、尿中に検出される病である。PKUは、発達遅延、精神遅延、湿疹、色素欠乏など多くの症状を小児に引き起こし、フェニルアラニン制限食を用いた食事療法など早期の治療を必要とする遺伝病の一つである。この他にも多くのこうした例があるが、それらは、ひとつの遺伝子に起こる変化が種々の細胞や組織に広汎な影響を与える事実を教えてくれる。
雑種強勢 (ヘテロシス、heterosis) :メンデルの「顕性の法則」では、雑種の形質は顕性親のそれと等しくなるがそれを超えることはない。しかし、雑種がどちらの両親よりも優れた性質や機能を表すことがある。これは古くから知られた現象で、「植物雑種の実験」からわかるように、メンデルもこれに気付いていた。最もよく知られているヘテロシスの例は、雑種トウモロコシで見出されたものである。1908年、ニューヨーク州ロングアイランドのコールドスプリング・ハーバーにあるカーネギー研究所のジョージ・シャルは近交系(自殖系)間の雑種F1の生産力が大きく増強することを明らかにし、この興味深い現象をヘテロシスと名付けた(Shull, G.H. (1908) The composition of a field of maize. Journal of Heredity. os-4: 296-301.)。シャルは、さらにトウモロコシの生産力は雑種の両親間で近親交配の度合いを示す近交係数Fが低下するほど高まることを明らかにした。その後、多くの遺伝育種学研究者の努力、中でも細胞質ゲノムが関与する雄性不稔(細胞質雄性不稔)と呼ばれる現象の発見により一代雑種の作成が容易になり、収量を50%-100%も増加させるヘテロシスの力を利用することで、20世紀の中頃には合衆国のトウモロコシ生産力は大いに増大した。今日、私たちが日々食する野菜の多くはヘテロシスを利用した一代雑種品種である。イネやコムギなどの自殖性の作物では他殖性のトウモロコシほど大きなヘテロシス効果が認められないが、それでも無視できない効果が認められ、これを見込んだ生産力増強に向けて、世界中で研究が進められている。
ヘテロシスを説明する分子的機構は未だ不明のままであるが、現在3つの有力な仮説が提唱されている。すなわち、顕性説(顕性相補説)、超顕性説(単一遺伝子座超顕性説)とエピスタシス説である。しかし、これらは互いに背反的ではなく、3つ全てが協働で関与していると考えられている。現在では、DNAのメチル化、ヒストンのアセチル化やリン酸化、低分子RNAによるRNA干渉などのエピジェネティック因子の関与が有力視されている(Liu, J., Li, M., Zhang, Q., Wei X. and Huang X. (2019) Exploring molecular basis of heterosis for plant breeding. Journal of Integrative Plant Biology 62: 287-298.)。そう遠くない将来には、メンデル遺伝学の拡張であるこの重要な遺伝現象のカラクリが説明される日がきっと来るであろう。
自家不和合性 (self-incompatibility, SI):一つの花器官に雌雄の生殖器を併せ持つ両性花や一つの個体で雌花と雄花を持つ雌雄異花では、自家受精(受粉)による同系交配を避けるために自他の花粉を識別する能力である自家不和合性を進化させたものが多い。すなわち、自分と同じ花粉は拒絶し自分以外の花粉との間でだけ受精が起こる。これは植物における自他認識の代表的なシステムであり、一つの遺伝子座(S遺伝子座)に存在する複数のアレルによって決まっているが、うえで述べた共顕性とは異なり、可変的あるいは可塑的な顕性・潜性関係が認められる。S遺伝子は雄蕊の花粉と雌蕊の柱頭や花柱で特異的に発現し、花粉細胞と雌蕊細胞の遺伝子型が一致する場合に不和合となる。従って、受精によって生じた植物のS遺伝子座は必ずヘテロ接合である。特に、雌雄の花器官が同一である同型花型の自家不和合性には配偶体型の(配偶子の遺伝子型で決定される)自家不和合性と胞子体型の(胞子体の遺伝子型で決定される)自家不和合性の二つがあり、どちらも単数体(半数体)の花粉(配偶子)が持つS遺伝子座におけるアレル間の相互作用が関与している。ナス科やバラ科の植物が示す配偶体型の自不和合性では、単数体(半数体)の花粉(配偶子)に存在する一つのSアレルが直接に表現型として現れる。一方、胞子体型のアブラナ科の植物では親植物体が持つ二つのSアレルの顕性・潜性関係によって花粉の表現型が決まる。特にアブラナ科の自家不和合性を決めるS遺伝子座には数多くのアレルが存在し、それらの間には顕性・潜性に関する複雑な階層構造が認められる(Yasiusa, S,. Wada, Y., Kakizaki, T., Tarutani, Y, Miura-Uno, E., Murase, K., Fujii, S., Hioki, T., Shimoda, T., Takada, Y., Shiba, H., Takasa-Yasuda, T., Suzuki, G., Watanabe, M. and Takayama, S. (2016) A complex dominance hierarchy is controlled by polymorphism of small RNAs and their targets. Nature Plants 3: 16206. DOI: 10.1038/nplants.2016.206)。
エピスタシス(epistasis): 第3章で述べたように、エピスタシスはメンデルの法則の再発見者の一人であったウィリアム・ベイトソンが1909年に発見して名付けた遺伝現象であり、広義には、複数の遺伝子座にある複数の遺伝子間の相互作用が同一形質に関与する現象である。メンデルはこの現象に気付いていたようであるが、生物が示す様々な表現型はメンデルが明示したような単一遺伝子によって決定されている場合はそれほど多くなく、むしろ例外である。多くの形質は複数の遺伝子座に存在する複数の対立遺伝子の相互作用によって支配されており、さらに多くの環境要因がこれに影響を与えている複雑な形質である。
ベイトソンが提唱したエピスタシスによれば、一つの生化学反応の異なる段階に影響を与える複数の遺伝子が関与している場合には、メンデルの観察した典型的な分離比は見られないことになる。例えば、2つの遺伝子が関わる単純な二因子交配の例では、F2世代の表現型の分離比が9:3:3:1ではなく、12:3:1、13:3、9:4:3, 9:7, 15:1などとなる場合がある。メンデルは実際に15:1の分離比をインゲンの花色で観察していた(第3章参照)。こうした期待比からのずれは、一つの遺伝子座の効果が他の遺伝子座の効果に影響を与えることから起こると考えられる。
一般に、二重変異体は遺伝解析にとって極めて有益な素材である。今、対照的な表現型を示す2つの遺伝子座の対立遺伝子が共に変異型である二重変異体で、一方の変異形質が他方の変異形質を覆い隠す場合、そのような遺伝子は上位にある(epistatic)と云い、逆に変異形質が覆い隠される遺伝子を下位にある(hypostatic)と云う。例えば、生物の発生において働く情報伝達のスイッチ調節経路,すなわち,その経路内の各構成要素が直線状に連なる他の構成要素を活性化あるいは阻害する(関与スイッチをオンにするかオフにする)場合には、そこで働く遺伝子の変異体間でエピスタシスが見られるとき、上位の遺伝子産物は下位の遺伝子産物に対して下流で機能すると結論できる。一方、ジョージ・ビードルとエドワード・テータムがアカパンカビで明らかにし、1958年のノーベル生理学・医学賞受賞の対象となった、連続した酵素反応による生合成や分解などの代謝経路や発生において特定の細胞グループを選択する経路などの基質依存経路(ポール・バーグ、マクシン・シンガー『ジョージ・ビードル“非凡な農民”:20世期遺伝学の誕生』、中村千春訳、)では、スイッチ調節経路の場合とは逆に、上位の遺伝子産物は下位の遺伝子産物に対して上流で機能する。エピスタシスの原理を用いた解析は遺伝子の発現調節経路を明らかにするうえの強力で重要な手法のひとつであるが、それだけでは必ずしも十分ではなく、遺伝子が発現する細胞・組織やタイミングの解析、遺伝子産物の機能解析などの総合的な解析が必要である。
伴性遺伝(sex-linked inheritance):性に伴う、より正確に言えば性染色体に伴う遺伝形式を伴性遺伝と呼ぶ。伴性遺伝は、1910年にコロンビア大学のトーマス・ハント・モーガンによる、ショウジョウバエの性染色体の一つX染色体に潜性白眼変異を支配する対立遺伝子(w, white)が存在するという発見を導く基礎となった遺伝現象であった。これを契機にしてモーガンたちは、特定遺伝子は特定染色体上にあるとする“遺伝の染色体説”を確立することができた(Morgan, T.H., Sturtevant, A.H., Muller, H.J. and Bridges, C.B. (1915). The Mechanism of Mendelian Heredity. New York: Henry Holt.)。ショウジョウバエでは、雌は両親に由来するX染色体を二本持つが、雄は雌親に由来するX染色体1本と雄に由来するより小さいY染色体1本を受け継ぐ。X染色体とY染色体とはほぼ非相同(遺伝構成が異なる状態)であるうえ、Y染色体上には性を決定する遺伝子以外にほとんど遺伝子が存在しないから、雄では母親から伝わるX染色体上の白眼遺伝子はヘミ接合の状態にあって赤眼を決める顕性対立遺伝子を持たない。メンデルの遺伝法則では、一つの遺伝子座には同じあるいは異なる二つの対立遺伝子が存在するとされたが、伴性遺伝では一方の性は対立遺伝子を欠いたヘミ接合の状態にあることになる。
伴性遺伝の例は人間でも多く知られている。女性はX染色体を二本持ち、男性はX染色体とY染色体を一本ずつ持っている。男性を決めているのはY染色体上の性決定遺伝子SRY(Sex-determining region Y)である。よく見られる伴性遺伝の例には緑色と赤色を識別できない赤緑色覚異常がある(色盲と呼ばれていたこの症状は、現在では色覚異常あるいは色覚多様性と呼ばれる)。人の色覚は網膜の錐体細胞に存在する3つの光感受性タンパク質色素のロドプシンによって媒介される。光の3原色は青、赤、緑であり、青を感知する色素の遺伝子は7番染色体にあり、赤と緑を感知する色素の遺伝子はX染色体に存在している。したがって、男性では一本のX染色体上の色素遺伝子に異常があればそのまま赤緑色覚異常として発現するが、女性ではホモ接合でのみ発現し、ヘテロ接合では保因者となる。男性のおよそ5%が何らかの形で赤と緑の識別に困難さを抱えているようである。
X染色体の不活化(X-inactivation):XX(雌)- XY(雄)型の性決定を行う生物種では、雌と雄でX染色体の量に差が生じるが、このような生物ではX染色体上の遺伝子量に補正が行われる。哺乳類の雌では、2本のX染色体のどちらか1本のX染色体が凝集して不活化する(ヘテロクロマチン化する)ことで、遺伝子量を補正している。三毛猫のまだら模様(白、茶、黒)はX染色体不活化のよい例である。猫のX染色体には毛の色を決める対立遺伝子が二つあり、Bは黒色(ブラック)を、Oは茶(オレンジ)色を決めている。雄はX染色体を1本しかもたないので黒か茶のどちらかとなる。一方、雌は2本のX染色体がBとOをヘテロ接合の状態でもつ場合、一方が不活化する細胞系譜(ある細胞を起源として細胞分裂を通じて生じた細胞集団)では黒か茶のパッチ模様が現れる。さらに、常染色体(XY以外の染色体)には色素形成を抑制する遺伝子Sが存在し、これが発現すると白色となる。この遺伝子は特定の細胞系譜でのみ発現し、そのような領域では白いパッチ模様が生じる。遺伝子Sの発現を決める仕組みは不明であるが、こうしてB, O, Sの3つの遺伝子の発現が絡んで黒、茶、白の三色を持つ三毛猫となる。従って、三毛猫は全て雌である。しかし、これにも例外があり、雄の三毛猫が生まれる場合がある。雌親のX染色体が減数分裂時に両極に別れず、一つの同じ娘生殖細胞へ入った場合には(染色体の不分離)、X染色体を2本もつ卵細胞が生じ、これがY染色体を持つ精子と受精すればXXYの染色体構成を持つ雄が生まれ、雄性の三毛猫となることがある。
染色体不分離は我々人間でもある頻度で起こる。性染色体が関与する場合はXXY、XXX、XO(Y染色体を欠く)などの染色体異常(異数体)の原因となる。XXYはクラインフェルター症候群と呼ばれる男性、XXXはトリプルXと呼ばれる女性、XOはターナー症候群と呼ばれる女性で、それぞれ特徴的な症状を表す。その他、常染色体に不分離が生じると1本過剰な染色体を持つ異数体(トリソミー、三染色体性個体)が産まれる。染色体が一本少ない異数体(モノソミー)はもちろんトリソミーでは例外を除いてほぼ全てが死産となる。トリソミーで生存可能な例外は、21番染色体(ダウン症候群)、18番染色体(エドワード症候群)と13番染色体(パトー症候群)で、特徴的で様々な症状を持つ赤ん坊が産まれ育つ。
倍数性(polyploidy):倍数性は配偶子に含まれる一組の染色体セットの正確な倍数的変化で生じ、植物で多く見られる現象である。倍数性には同質(auto)倍数性と異質(allo)倍数性がある。同質倍数性は同じ染色体構成をもつゲノム全体の整数倍変化を云う。遺伝子の相加的効果で細胞や個体のサイズが2倍体よりもずっと大きくなるが、染色体の不均等な分配が行われて稔性が低下し、子孫を得ることが難しい。しかしそれも種芋で繁殖するジャガイモなどでは稔性は問題とはならず、高収量は有利である。同質倍数性の好例としては他に、ゲノムの概念を広めた京都大学の木原均教授が開発した種無しスイカがあげられる。種無しスイカは、イヌサフランの種子や球根に含まれるアルカロイドであるコルヒチンで減数分裂時の紡錘体(染色体と両極を結ぶ構造)の形成を阻害し、それにより染色体数を倍加した4倍体と通常の2倍体の交配から育成した3倍体である。染色体セットを3組持つ3倍体は減数分裂の異常で種がほとんどつかないので食べたりジュースにしたりするのに便利だから、今でも東南アジアでは人気が高い。しかしなぜか、日本では一時ほどの人気はないようである。
一方、異質倍数性は2種類以上の異なったゲノムをもつ倍数体を云う。近縁種間で形成される場合が多く、減数分裂では同祖染色体(起源を同じする相同な染色体)の対合が抑制され、染色体が通常の2倍体のように振る舞う複二倍体化の結果、稔性を持つ倍数体として増殖可能である。異質倍数体の代表例はコムギである(これについては第3章を参照)。
単為生殖(parthenogenesis)・無配偶生殖(アポミクシス、apomixis):単為生殖は単為発生とも呼ばれ、雌あるいは雄が単独で発生し子が産まれることを云う。卵から雌のみが産まれる産雌性単為生殖はセイヨウタンポポ、メンデルが交配実験に用いたヤナギタンポポ属(第5章と7章参照)や害虫として知られるアブラムシで見られる繁殖様式である。一方、産雄性単為生殖はミツバチ、アリなど膜翅目昆虫やダニなどで見られる。ミツバチはメンデルが交配実験を手がけた昆虫で(第7章参照)、アリなどと同じく有性生殖と単為生殖の双方を行う。これらの社会性昆虫は、女王バチのみが生殖に預かり、受精卵からは雌が産まれ働きバチとなるが、受精を経ずに半数性の卵がそのまま発生した場合には雄バチが産まれる。女王バチは雄バチとの交配で得た精子を受精嚢に蓄え、産卵時に受精を制御して雄雌の産み分けを行っている。このように、受精卵から発生した2倍体の雌と未受精卵から発生した単数体の雄のスイッチが起こる繁殖様式は単数二倍体性(haplodiploidy)と呼ばれている。雌バチは女王バチにも働きバチにもなれる遺伝構成を持つが、女王バチになるのはロイヤルゼリーという特殊な餌を与えられた雌バチのみである。なお、膜翅目では、父由来の遺伝子は同一だから、同一の母から産まれた姉妹の遺伝的な近縁度は、通常の1/2ではなく3/4で、父母を共有する姉妹の血縁の濃さは、雌雄を問わず自分の子供に対する血縁の濃さを上回ることになる。これが、姉妹が自分の子供よりも女王を懸命に養う理由になっている。
植物の単為生殖はアポミクシスと呼ばれ、これも受精を伴わない無性的な繁殖様式である。球根やムカゴはこれに含まれるが、挿木や接木による繁殖はこれに含めない。アポミクシスには卵細胞が発生を始める場合とともに花粉の精細胞が発生を始める場合があり、どちらでも生じる子孫は遺伝的に同一なクローンである。植物の繁殖戦略として多くの様式が存在するが、メンデルが実験材料に用いたヤナギタンポポ属では2倍性の雌性配偶子が単為発生する複相胞子生殖(diplospory、ディプロスポリー)と呼ばれる様式が働いている(メンデルを悩ませたヤナギタンポポ属のアポミクシスについては、第5章と7章を参照)。
細胞質遺伝(cytoplasmic inheritance):メンデルの遺伝法則の重要な例外事例として新たな研究領域を生み出した細胞質遺伝あるいは母性遺伝と呼ばれる遺伝現象は、メンデルの遺伝法則の再発見者の一人であったカール・コレンスによって1909年に報告された(Rheinberger, H. J. (2000). "Mendelian inheritance in Germany between 1900 and 1910. The case of Carl Correns (1864-1933)". Comptes Rendus de l'académie des Sciences, Série III. 323: 1089-96.)。コレンスはオシロイバナ(Mirabilis jalapa)の変化に富んだ葉色について研究した。メンデルの法則では、形質は雌雄の性別に依らずそれとは無関係に振る舞う、すなわち正逆交配で同一の結果をもたらすのに、オシロイバナの斑入り現象では違っていた。オシロイバナには緑色の葉、黄色の葉、それと斑入りの葉をつける3種類の植物がある。これら3種類の植物から得た花粉をそれぞれ同じ植物の別の枝あるいは別の植物の雌蕊に受粉させると、子孫の表現型は花粉親がどれであっても全て母親(種子親)の表現型と同じであった。例えば、緑色の個体の柱頭を緑色の個体の花粉で受粉させると緑色の子孫が、あるいは斑入りの柱頭を斑入りの花粉で受粉させると斑入りの子孫が生じた。ところで、斑入りの柱頭を緑色の花粉で受粉させると斑入りの子孫が生じたが、緑色の柱頭を斑入りの花粉で受粉させると子孫は緑色であった。すなわち、子孫の表現型は全て母親(柱頭の提供親)の表現型と同じであった。コレンスの発見した細胞質遺伝は細胞質に存在する細胞小器官である葉緑体(chloroplast)の突然変異によるものであることが後にわかった。
1952年にはミッチェルによってカパンカビで(Mitchell, M. B., Mitchell, H. K.(1952) A case of “maternal” inheritance in Neurospora crassa. Proc. Natl. Acad. Sci. USA 38: 442-449.)、翌1953年にはボリス・エフルッシらによって酵母(イースト)で(Ephrussi, B. 1953. Nucleo-cytoplasmic relations in micro-organisms: their bearing on cell heredity and differentiation. Oxford.)多くの変異体が見出された。これらの変異体はどれも別の細胞小器官であるミトコンドリア(mitochondoria)の変異体であった。現在では光合成と呼吸をそれぞれ担うこれらの細胞小器官には固有のゲノムが存在し、細胞質遺伝の原因であることが分かっている(Berg, P. and Singer ,M. (2003) George Beadle, An Uncommon Farmer: The Emergence of Genetics in the 20th Century, Cold Spring Harbor, New York.ポール・バーグ、マクシン・シンガ(2014)ジョージ・ビードル、非凡な農民:20世紀遺伝学の誕生、 中村千春訳訳)。ところで、葉緑体DNAは、1963年にルース・セイガーと石田政弘によってクラミドモナスで、1972年にはリチャード・コロドナーとへーマン・K・テワリによってエンドウで発見されたが、ミトコンドリアDNAについては、突然変異体に関する多くの研究がミトコンドリア遺伝子の存在を示唆し多くの生化学的な研究が続けられたにもかかわらず明確な証拠は得られなかった。しかし1964年に、ウィーン大学生化学研究所のエレン・ハッスルブルナーらによるショ糖密度勾配遠心分離法によって酵母のミトコンドリアDNAがついに捉えられた(Schatz, G., Halsbrunner ,E. and Tuppy, H. (1964) Deoxyribonucleic acid associated with yeast mitochondria. Biochem. Biophys. Res. Commun.15, 127-132.)。
 図2 リン・マーギュリス
図2 リン・マーギュリス
|
細胞小器官のDNAに関して生物学界に最も大きな影響を与えたのは、1966年のリン・マーギュリス(図2)による細胞内共生説であった。ボストン大学の女性研究者であったマーギュリスの投稿論文は、突拍子もないとして少なくとも15の科学雑誌から却下され、翌年になって漸く掲載された。細胞内共生説は、アルファ・プロテオバクテリアという真正細菌がある種の真核生物の母体となった細胞に共生して呼吸を担うミトコンドリアとなり、さらに原核細胞であるシアノバクテリウムの共生により光合成を担う葉緑体ができたとする(Margulis (Sagan), Lynn (1967). "On the Origin of Mitosing Cells". Journal of Theoretical Biology. 14 (3): 225-274. )。
細胞質遺伝については、細胞質ゲノムの遺伝的多様性とともにそれと核ゲノムとの相互作用の多様性を解析した京都大学農学部の常脇恒一郎教授グループの仕事が特筆される。木原均の研究を引き継いだ常脇は、1965年以来、共同研究者とともにコムギとその近縁属である野生エギロプス属46種の保有する細胞質ゲノムとパンコムギ12系統の核ゲノムを組み合わせた552系統もの核細胞質雑種(細胞質置換雑種あるいは核置換雑種とも呼ばれる)系統を作出し、それらが示す様々な形質のスペクトラムを解析して、コムギ・エギロプス属の核ゲノムと細胞質ゲノムの系譜および両者の共進化関係を明確にしている(Kihara, H. (1951) Substitution of nucleus and its effects on genome manifestations. Cytologia 16: 177-193.; Tsunewaki, K (2009) Plasmon analysis in the Triticum-Aegilops complex-Review. Breed Sci 59:455-470, and erratum (2010) 60:177-178. ; Tsunewaki, K., Mori, N., Takumi, S. (2019) Experimental Evolutionary studies on the geneticx autonomy of the cytoplasmic genome ‘plasmon’ in the Triticum (wheat)-Aegilops complex. PNAS 116 (no.8): 3082-3090.)。
ここで述べた例外的な現象の多くはメンデルの遺伝法則の拡張として説明できるものであり、法則の普遍性を些かも損ねるものではない。細胞質遺伝(母性遺伝)にしても、形質が遺伝子によって決まるという原則に変わりはない。一般に、法則や原理が本質的であればあるほどその敷衍するところは広いと考えられるが、こうした例外の存在もそのことをよく示している。遺伝形質は遺伝子が担うとしたメンデルの遺伝法則があったからこそ、その拡張があったのである。
メンデルの遺伝法則の意義
メンデルの遺伝法則の最大の意義は、子孫に伝達するのは個別の実体としての遺伝子であって形質はその結果であるという事実を明らかにしたことであった。遺伝子が形質を決めるという「粒子遺伝(particulate inheritance)」の概念はメンデル遺伝学の本質である。表面に表れる形質とそれを支配する遺伝子との因果律を明確にし、一つの形質には一つの遺伝子が対応するとしたメンデルは、これにより遺伝現象(子孫への形質の伝達)の理解に新たな飛躍をもたらし、その後の遺伝学発展の礎を築いた。メンデルの叡智の結晶としての遺伝法則の発見は、後に、トーマス・ハント・モーガンのグループが1910年代にショウジョウバエを材料に実施した一連の実験により明らかにした、特定遺伝子は特定染色体上の特定の場に存在するとした「遺伝の染色体説」および、一つの遺伝子は一つの酵素に対応するとしてジョージ・ビードルとエドワード・テータムが1941年に発表した「一遺伝子一酵素説」に引き継がれてゆく。「遺伝の染色体説」は、遺伝子が染色体上に直線的に配列していること、個々の遺伝子間の相対的な距離はそれらの組換え頻度によって測定することができ、従って遺伝子の並びと互いの距離に基づく遺伝地図(連鎖地図、染色体地図)が描けることを明らかにした。これによって、形質を次代に伝達する役割を担うメンデルの遺伝子は、染色体上の実体として捉えられることになった。メンデルが魁となって門戸を拓いた遺伝学をさらに発展させ、「遺伝の染色体説」を確立した貢献により、モーガンは1933年にノーベル生理学・医学賞を受賞している。さらに、メンデルの遺伝子の本質的な機能が酵素の合成指令であることを決定的にしたビードルとテータムの「一遺伝子一酵素説」は遺伝学に画期的な発展をもたらした。ひとつの酵素が複数のポリペプチド(アミノ酸が連続的に並んだタンパク質)で構成される場合があることから、現在では、この説は「一遺伝子一ポリペプチド説」と呼ばれる。二人は1958年のノーベル生理学・医学賞を受賞している。こうした古典遺伝学の潮流はさらに勢いを増し、ついには1953年のジェームス・D・ワトソンとフランシス・H・クリックによるDNA二重らせんの発見を導くことになる。二人は1962年にノーベル生理学・医学賞を受賞している。(Watson, J.D. and Crick, F.H. (1953). "A structure for deoxyribose nucleic acids" (PDF). Nature. 171 (4356): 737-738.; Orel, V.(1996) Gregor Mendel: the first geneticist. Oxford University Press, Oxfordshire,UK.; Hartl, D. and Orel, V.(1992) What did Gregor Mendel think he discovered? Crow, J. F. and Dove, W.F.(eds.) Perspectives: Anecdotal, historical and critical commentaries on genetics. Genetics 131: 245-253. ; ジャン・ドゥーシュ(2015)進化する遺伝子概念 佐藤直樹訳 みすず書房)。
第2章ですでに見たように、メンデル以前に考えられていた遺伝様式は混合遺伝あるいは融合遺伝(blending inheritance)であった。「混合遺伝」は科学的な検証が行われたことのない、いわば机上の論だったが、19世紀半ばまでは遺伝に関する考え方の主流であった。「混合遺伝」に基づけば、種内に存在する変異あるいは多様性は世代ごとに薄まって行き、遂には均一な純系ともいうべき状態になるはずである。これは、当時の博物学者・生物学者や育種研究者が解析の対象にした形質の多くが、複数の遺伝子が関与する、しかも環境に大きく影響される量的形質であって、人為交配によって生じる雑種の形質は一般に両親の中間型に近づくという観察を根拠にした考え方であった。この考え方の最大の難点は、遺伝は混じり合っても実体を失うことのない個別粒子ではなく、むしろいわば液体のような物質によって担われており、受精により一旦混じり合えば再び分けることはできないとした点であった。「混合遺伝」ではいったん隠れた親の性質が後代で再び現れるという復帰あるいは「隔世遺伝」の現象を説明できない。例えば、家系を通じて何代も現れなかった青い目とブロンドの髪をもつ子供が茶色の目と茶色の髪をもつ両親から生まれることがあるが、これを説明できない。メンデルの「粒子遺伝」の重要性は、遺伝情報を担う実体が粒子的な性質を持ち、受精によって合一した後でも変化することなく子孫に伝えられること、さらに子孫では一定の割合で両親型が表れるという事実を明らかにしたことにあった。メンデルの「粒子遺伝」説によって「混合遺伝」は明確に否定された。なお、チャールズ・ダーウィンの従兄弟で、「優生学」の創始者であったフランシス・ゴルトンは、子は両親から半分ずつ、祖父母から4分の1ずつ、さらに相祖父母から8分の1ずつの形質を受け取るとする「祖先遺伝の法則」によって「隔世遺伝」を説明した。実は、多くの遺伝子が関与する量的形質では、混合説によっても粒子説によっても、種内の変異は世代ごとに狭まって、遂には両親の平均値に近づき均一な状態になる。メンデルの「粒子遺伝」と「混合遺伝」の論争については第6章で改めて見ることにする。
メンデルが発見した遺伝法則の第2の意義は、遺伝子の担い手が雌雄の配偶子であることを明確にし、さらに配偶子を通じて遺伝子が、従って形質が次代に伝達する仕組みには数学的な規則性があることを明らかにしたことであった。分離の法則は、形質の分離が雌雄の配偶子によって運ばれる対立遺伝子の無作為な組み合わせの結果であることを、さらに雌雄の配偶子の受精における貢献が等価であることを前提として成り立つ。メンデルは、先人たちの報告と予備実験を含むエンドウの交配実験で得た結果から、このことを確信したに違いない。科学は、ある概念を事実によって裏付ける行為、すなわち組み立てた作業仮説を実験によって実証する行為であるが、メンデルはこれを忠実に実行した。「植物雑種の実験」からは、メンデルが形質分離の基盤は対立遺伝子の無作為な分離と組み合わせであることを見抜き、これを実証するために、計画に従って丹念な交配実験を続けたことが明瞭に見て取れる。
一方、「種の起原」で壮大な進化の理論を打ち立てたダーウィンは自ら「パンジェネシス(パンゲン説)」と名付けた説で遺伝の現象を説明していた。「パンジェネシス」では、体の各部を構成する細胞がそれらの細胞の特徴を決めている個別の粒子ジェミュールを持っており、いったん生殖細胞に集まったジェミュールが再び体全体に拡散して各部を構成する新たな細胞を生みだすと説明された。ダーウィンはラマルクの獲得形質の遺伝を支持し、自然選択によって獲得された有利な性質が子孫に伝わると主張したが、「パンジェネシス」はこれを説明するために考え出された説で、細胞学的に明らかな誤りを含んでいた。体を構成する細胞は全てが単一の受精卵から派生する。一方メンデルは、形質の遺伝が対立遺伝子の配偶子への分配と、雌雄配偶子の受精を通じた再結合によること、従って交配によって変異が加わったり失われたりすることはないと主張し、「パンジェネシス」を批判した。メンデルは、しかし論文中で直接にはダーウィンの「パンジェネシス」に言及していない。この点については第5章と6章で論じることにする。
メンデルの成功の秘訣
メンデルが遺伝の法則を発見できた理由、すなわち成功の理由あるいは秘訣を考えてみる。メンデルの研究結果を解説した書物や論文では概ね次の5つの理由が挙げられている(Orel, V.(1996) Gregor Mendel: the first geneticist. Oxford University Press, Oxfordshire,UK.; Hartl, D. and Orel, V.(1992) What did Gregor Mendel think he discovered? Crow, J. F. and Dove, W.F.(eds.) Perspectives: Anecdotal, historical and critical commentaries on genetics. Genetics 131: 245-253. ; ジャン・ドゥーシュ(2015)進化する遺伝子概念; 佐藤直樹訳、みすず書房; Iltis, H.(1932) Life of Mendel, Eden and Cedar Paul(英語版翻訳), London, George Allen & Unwin Ltd.)。
1)適切な材料を選んだ
エンドウはマメ科のエンドウ属(Pisum)に属する。エンドウは、ヨーロッパでは主にスープの具材として食される園芸作物である。圃場でも温室でも育て易く、根粒細菌による空中窒素固定のお陰で窒素肥料の施肥に悩まされる必要がなく、耐寒性に優れ、種子を植えてから次代の種子を得るまでの世代期間が55日から75日と比較的短く、年に少なくとも2世代を得ることが可能である。加えて、多くの顕花植物と同じように雌雄両性で同一の花器官に雄蕊と雌蕊が存在するうえ、自家受精により繁殖するから純系が得やすい。実際には、他の自殖植物と同じく稀に他個体の花粉による受精が起こることがあるから、これを避けて自家受精を確実にするために、母親植物体の個々の花器官に袋を掛けて望まない受粉を避ける必要がある。人為交配も比較的容易で、雄蕊を除くための除雄の操作とともに、花粉親に選んだ他個体から集めた花粉による雌蕊への授粉操作で、雑種を作ることができる(こうした除雄と人為的な授粉による雑種の作製法はケールロイターによって開発された)。
さらに1回の授粉でひと莢に6から9個ほどの種子を着粒するなど実験材料として好都合な多くの性質を備えている。特に遺伝解析では多数の子孫を扱う必要があるから、短い世代時間とともに植物では授粉により多くの種子を着ける性質は重要である。雌穂(雌花)への雄穂(雄花)による1回の授粉で1穂に600粒もの種子を着けるトウモロコシには遠く及ばないが、小花への1回の授粉で1個の種子しか着けないコムギやイネに比べればエンドウは遺伝育種操作のうえで格段に便利な実験材料であることが分かる。
2)対照的で安定した形質を解析の対象に選んだ
メンデルが最終的に選んだ形質は種子に関するものが2種類、植物体に関する形質が4種類、種子と植物体の両方に関する形質が1種類だったが、どれも肉眼で明確に識別が可能で、比較的安定した定性的な形質だった(茎の長さすなわ草丈は定量的な性質であるが、メンデルは一定の判定基準を設定して、これを定性的な性質として扱っている。第3章参照)。それでもなおメンデルは、形質評価と判定に際して慎重だった。メンデルが論文で繰り返し述べているように、これらの定性的な形質も天候や病害虫などを含む栽培環境要因によって判定基準からずれることがしばしばで、メンデルは形質評価に細心の注意を払わなければならなかった。形質評価は今でも遺伝学や育種学の研究者を悩ます難題で、その正確な判定は決定的に重要である。「植物雑種の実験」では、例えば次の記述に見られるように、判定の際の苦労の跡が随所に記されている。「交配には、同一系統の多数の植物から、もっとも健全な個体だけを選んだ。弱い株では、すでに雑種の初代で、その後代ではなおのこと、多くの子孫が開花しないか、開花しても少数の不良種子ができるだけで、常に確実な結果が得られるとは限らないからである」、「子葉の色については、緑色の発達が不良で浅黄色に見えることがある。この点は雑種植物の特徴ではなく親植物でも同様の傾向が見られるが、それは当該個体にのみ限られた傾向で子孫に安定的に伝わる形質ではない」、「発育の途中で昆虫による加害を受けた種子は色や形態が変わることがあるので、選別に習熟して誤りを避けなければならない」、「莢はその形と内部の種子の色が完全に発達し成熟して乾燥するまで植物体上につけておかなければならない」。その上でメンデルは、2年間で実施した予備試験の結果から22系統の品種群をえらび、最終的に7種類の定性的な性質を備えた系統を交配実験の材料に選んでいる。
3)純系間の交配を行った
この点は2)と密接に関係する。問題とする形質が代々安定して子孫に伝わることを確認するためには、自殖(自家授粉)後代でその形質が確実に全ての子に伝わることが明らかでなければならない。自殖(自家受精)後代への伝達が不安定であれば、解析の対象とする形質の雑種後代における正確な動向を探ることは難しい。多くの育種家や園芸家が失敗したのは、いくつもの遺伝子が関与する複雑な形質を対象としたからだけでなく、純系を交配の出発材料としなかったからであろう。メンデルは、自家受粉を繰り返し、毎代同じ形質を安定して表す純系をまず作出し、それらを交配に供した。識別可能な特定形質について純系を用いるのは、今では遺伝解析のための必須手段である。メンデルの遺伝法則が再発見された3年後の1903年にデンマークの遺伝学者ウィルヘルム・ヨハンセンが、生物の集団は純系に達すると選抜を加えてもそれ以上変化しないとする純系説を提唱した。ヨハンセンは市場から買って来たインゲンマメ(Phaseolus vulgaris)の種子の重さを測定し、それが統計的に正規分布を示すことを確認した後、それらを幾つかの階級に別けて、それぞれの階級の中で自家受粉により得た各子孫を調査した。すると各階級の種子重の平均値は原品種のそれと異なり、重い種子からは重い種子が、軽い種子からは軽い種子が得られた。ヨハンセンはさらに原品種を出発点として自家受精により6世代に渡って、それぞれ重い種子と軽い種子を選んで継代し19系統を育てた。すると各系統内では種子重の値の平均値は変化せず正規分布を示し、各系統はそれぞれ純系に達したと結論した。ヨハンセンはこの結果から、生物集団が純系に達すればダーウィンの選択はもはや無効であるとする純系説を提唱することで遺伝学に重要な基礎的貢献をした。ヨハンセンの純系説は、メンデルがエンドウで明らかにした安定親型の出現に関する説明を支持するものであった。
4)交配の子孫に現れる形質を数量的に解析した
メンデルが論文中で明確に指摘したように、解析の対象となる対立形質の数が増すにつれて子孫で生じる形質の組み合わせの数は一定の規則にしたがって急速に増加する。解析の対象とする形質の数を限定するか、解析する個体の数を十分大きくしなければ、雑種の子孫に伝達される様式に潜む規則性を見いだすことは難しいだろう。形質が全て独立であるとすれば、両親で異なる形質の数がn対ある時、子孫で生じる組み合わせの数は3n、組み合わせ全体に属する個体の数は4nとなる。従って、解析に要求される個体数は幾何級数的に大きくなる。ウィーン大学で、物理学教授のヨハン・クリスチャン・ドップラーと数学教授のアンドレア・ヴァン・エッティングスハウゼンから確率論と組み合わせ理論を学んでいたメンデルは、特定の因子の分配を明らかにするためには雑種の子孫で現れる両親型(ホモ接合型)と雑種型(ヘテロ接合型)の出現数を正確に数える必要があることを正確に理解していた。その上で、F2における形質分離の比率がF1における因子(対立遺伝子)の分離比に基づくとする結論を下した。メンデルの発見した「独立分離の法則」に従えば、これはn対の相同染色体上にあるn対の対立遺伝子が関与する交配にそのまま拡張出来る。雑種後代の形質の分離とその基礎になる対立遺伝子の分離に関するこの単純で美しい数学的関係を明らかにした点でメンデルは先人達を超越していた。なおメンデルは、連鎖(同一染色体上に複数の遺伝子が並んで存在すること)に言及していないが、この点については第6章で述べる。
5)科学的方法論を採用した
研究に当って科学者が科学的な方法論を採用するのは当然のことで、この指摘は自明のことと思えるだろう。知識を獲得するための科学的方法論が生まれたのは17世紀に入ってからであった。科学的方法論の要素を段階的にまとめれば一般に次のようになる。課題の設定:意味のある課題や質問を解答可能と考えられる形で提示する。情報の収集:課題の解決にふさわしい情報を集める。仮説の設定:課題への解答を検証可能な仮説の形で提示する。実証実験の実施:仮説の確からしさを実験や観察によって検証する。議論と修正:仮説が棄却できないところまで修正を繰り返し、矛盾のない科学的理論として構築する。もちろん、反証の可能生も検討する必要がある(カール・ポパー著(1978)果てしなき探求―知的自伝、森博訳、岩波現代選書)。通常、科学論文は、序論(Introduction)、材料と方法(Materials and Methods)、結果(Results)と(And)考察・議論(Discussion)の頭文字をとって、IMRADと呼ばれる構成で記述される(Day, R.A. (1988) How to Write and Publish a Scientific Paper. 3rd Edition. ORYX PRESS.)。メンデルが「植物雑種の実験」で採用した方法は、当時の生物学者あるいは博物学者達が一般的に依拠した方法論、すなわち情報収集、分類整理と観察に基づく記載という伝統的な博物学的方法論に留まるのではなく、まさに現代的な科学的方法論であった。メンデルは、一般的・普遍的な規則を実験データから導き、その規則の確からしさを実験事実によって検証し、さらにその規則から導かれる新たな事実の実験による検証を試みる帰納法と演繹法を組み合わせた科学的方法論を採用した。メンデルの論文の書き進め方からは、メンデルが予め遺伝法則を念頭に置いた上で、当然導きだされる結果を予想して実験を組み立てたようにも見える。メンデルは確信を持って実験を計画し、着実に実験を積み重ね、質的にも量的にも検証に耐えうる十分なデータを得て結論を下したのであった。
メンデルはこうして遺伝の法則を発見した。全ての成功に共通するであろうが、メンデルの成功にはしかるべき理由があった。だが、この発見が「メンデルの遺伝法則」として「再発見」され、科学界で評価されるまでにはその後35年という歳月が必要だった。さらに、「メンデルの遺伝法則」が生物学上の普遍的真理として認められ不動の地位を獲得するまでには、「再発見」後にもなお20年以上という時間がかかった。それまでの常識を超えた概念が世に受け入れられるには、それを受け入れるだけの土壌が必要である。時代に先駆けた優れた仕事が本当のブレイクスルーとなるには一定の時間がかかる。これはあらゆる事象と場面を通じた真理であろう。
| <<前の章 |
TOP |
次の章>> |
|
Copyright (C)
2021 Chiharu Nakamura All Rights Reserved.
|
|
|