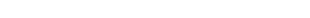研究の紹介
研究者探訪

生物の細胞膜には様々な膜タンパク質が存在し、連携して動くことで機能を発揮しています。膜タンパクや分子はどのように相互作用し、構造を変化させてその機能を発揮しているのでしょうか。今回は膜タンパクの構造解析を中心に研究を行われている竹田先生にお話しを伺いました。
聞き手:先生のご専門は「構造生物学」といわれる、生体分子や複合体の姿かたちを明らかにすることで生命現象の謎を解く研究分野ですね。先生のご研究のスタート地点はどのようなものだったのでしょうか?
学生時代に配属された研究室で与えられた研究テーマは、イオンチャネルタンパク質という膜タンパク質に関してでした。膜タンパク質とは、我々の細胞の生体膜にあるタンパク質のことで、その中でもイオンチャネルタンパク質とは、特定のイオンを細胞膜の内外に透過させる役割を持つタンパク質です。細胞や細胞内小器官は、状況に応じてイオンチャンネルが膜内外のイオン濃度調整を行うことで様々な機能を得ています。私は「平面膜法」という、イオンが膜を通ると電流を検出する方法を使って、イオンチャネルの機能を電気生理学的に調べていました。しかし、この方法では「現象」を追うだけで、機能は分かっても動作原理を知ることはできません。動作原理を知るには、タンパク質の「構造」の情報を基にした議論が必要です。私は博士課程進学を機に、膜タンパク質の構造解析で世界的に有名な東京大学の濡木研に移り、マグネシウムイオン(Mg2+)チャネルについて研究を始めました。
カリウムやマグネシウムなどの金属イオンは、水中ではいくつかの水分子に囲われています。これまでの研究で、カリウムイオン(K+)チャネルタンパク質の構造が解明され、K+がチャネルタンパク質を透過するときには、囲っている水がはぎ取られた(脱水)状態であることが分かっていました。それにくらべMg2+は、非常に脱水しにくい性質を持っています。はたしてMg2+は、脱水した状態でチャネル内を透過しているのかどうか、当時は分かっていませんでした。私は博士課程時代に、この構造を解明し、Mg2+イオンは水和した状態のままチャネルを透過していることを明らかにしました(図1)。私はこの研究を通して、構造解析研究が持つ力、学術的な説得力の凄さを実感しました。
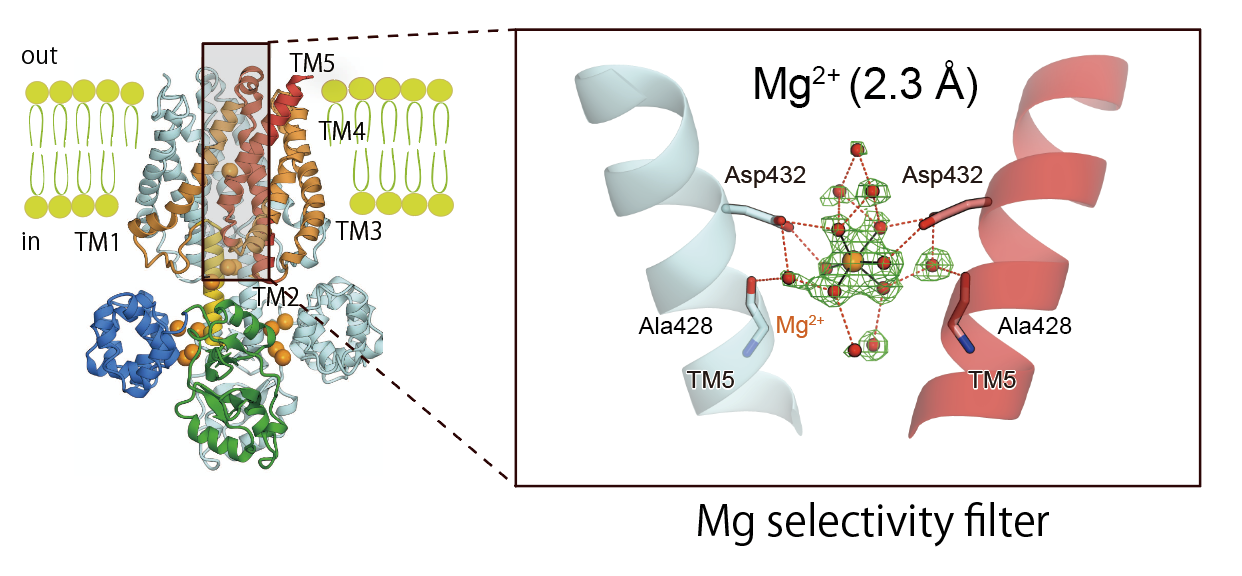
図1:左)マグネシウムイオンチャネルの構造 右)マグネシウムイオン(中央、オレンジ丸)が水分子(赤丸)を纏った水和状態のまま、チャネルタンパク質内を透過している様子。
聞き手:現在の先生の研究の中心は膜タンパク質の中でも、「膜挿入タンパク質」についてですね。
当時(2010~2015年)、構造解析の世界では、バクテリア由来の膜タンパク質の研究が主流で、あらゆる膜タンパク質の構造が解かれていた時代でした。バクテリアが持っている膜タンパク質は、それより高等なほとんどの生物種も持っていて、それほど構造も変わりません。新しい構造を発見したいなら、バクテリアが持っていない、真核生物のオルガネラ(小胞体、ミトコンドリア、ゴルジ体等)の膜タンパク質をターゲットにするのが良いと考えました。
ほとんどの膜タンパク質は、「イオンチャネル」や「トランスポータ」と言われる物質の輸送に関わるものです。しかし、私が面白いと思ったのは、「膜挿入タンパク質」でした。これはまだ完全な形に折れたたまっていない、未成熟の膜タンパク質(基質タンパク質)を認識して取り込み、それが正しい膜タンパク質の構造になるように整形して、膜内に挿入する機能を持っています(図2)。
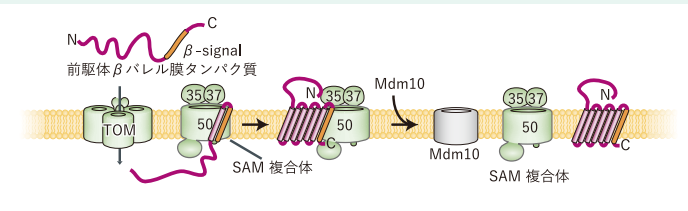 図2:膜挿入タンパク質の機能の概念図。
図2:膜挿入タンパク質の機能の概念図。
当時、その存在は分かっていましたが、どんな形で、どのように折れたたみを助ける(整形する)のかについては全く分かっていませんでした。この解明はおそらく非常に競争が激しくなるだろうけども、重要な膜タンパク質だと思い、研究対象として選びました。
聞き手:当時未開拓であったテーマを選ばれたわけですね。大変ではなかったですか?
当時、オルガネラ膜タンパク質、さらに膜挿入タンパク質を研究していた京都産業大学の遠藤研究室に移りました。遠藤研では膜挿入タンパク質「SAM複合体」の構造解析を行いましたが、苦労の連続で、論文が出るまでに5年くらいかかりました。
タンパク質の構造解析は研究というより「ものづくり」に近いです。通常、目的タンパク質は、出芽酵母に作らせて回収し、精製することで大量合成を行うのですが、これが全くうまくいきませんでした。こういう時は、それまでの定石から離れた、突拍子もない方法を持ってこないといけません。通常は、出芽酵母のプラスミドという環状DNAにターゲットタンパク質をコードする遺伝子を組み込んで、目的のタンパク質を発現させる、という方法が主流でしたが、私は直接出芽酵母のゲノムに遺伝子を入れてしまってタンパク質を増やすという方法を採用し、成功しました。次に、SAM複合体を精製しなくてはなりません。様々なタグを使った精製法を試しましたが、夾雑タンパク質が多く精製度が高くなりませんでした。そこで、これまでの大量精製にはほとんど用いられていない ProtA タグを用いることで、うまくSAM複合体を精製することができました。これらの方法はそれまでの構造解析の分野では前例のない方法でした。
しかし、タンパク質が採れたとしても、いざ構造を見るとなると、これが非常に難しいのです。私が用いたタンパク質の構造解析の手法は、急速凍結したタンパク質試料を電子顕微鏡で見る方法です。タンパク質の精製条件が悪かったり、タンパク質が壊れていたり、なかなかうまくいきませんでした。タンパク質試料を作っては試し、作っては試しを繰り返す苦労の連続でした。
「タンパク質の機能解析」研究では、研究目標の30%の結果でも、なんとかデータが出れば、論文にすることはできます。しかし構造解析は、構造がきちんと決定できなければ、論文になりません。構造がきれいに解けなければ、全部が無駄になる、いわば0か100かの世界です。ポスドク時代は非常に苦しい5年間を過ごしました。構造が解けたときは「やっと解けた」ではなく、「解放された」という感想でした。
聞き手:構造解析研究を行うには根気が必要なのですね。
そうですね。その一方で、構造解析研究の怖いところは、競争相手が世界中にいるという事です。まだ解かれていないタンパク質というのは、データベースを見れば一目瞭然なので、日本だけでなく、世界中の研究室のターゲットがそこに集中します。ですのでいかに早く構造を解くか、ということが非常に重要になります。
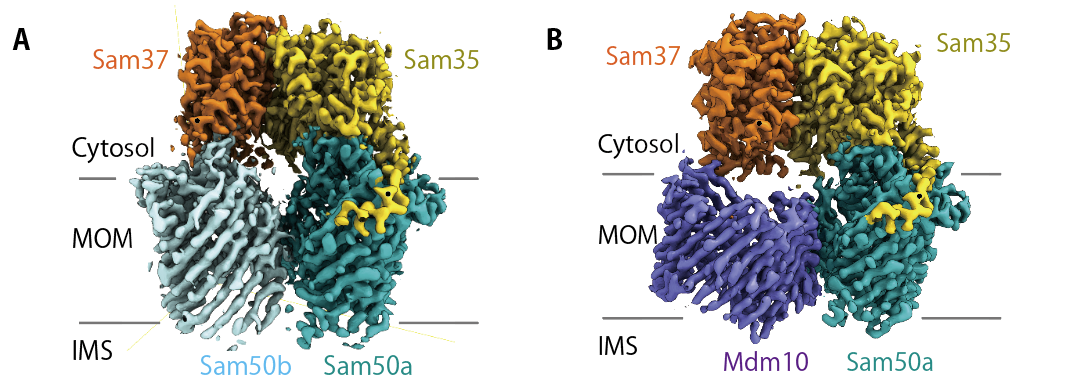
図3:SAM複合体 (A) 及びSAM-Mdm10複合体 (B) のクライオ電験構造
私はこのタンパク質の二種類の違う構造、SAM複合体と、SAM(mdm10)複合体を解きました(図3)。この二つの構造それぞれについて、ビッグジャーナルに論文が出せていたはずですが、その1か月前に、アメリカのチームによってSAM複合体の構造が解かれてしまいました。そのため、私は二つの構造を合わせて、ひとつの論文として出さざるを得ませんでした。そのくらい競争は激しく、過酷な世界です。構造解析の研究は、常に競争相手が何人居て、今どういう段階にあるのか、不安に駆られながら行わなくてはいけなくて、構造が解けたとしても一喜一憂できないところが有ります。
次に、苦労の末、SAM複合体と、基質タンパク質が折れたたみの中間構造で止まっている状態とで結合しているタンパク質の作成に成功し、構造を解くことが出来ました。その構造から、実際にSAM複合体が基質タンパク質をβバレルに整形する様子をこの目で見ることができました(図4)。その時はタンパク質の造形美、生命の神秘を感じ、興奮して1週間くらい寝られませんでした。このタンパク質に関して、長年苦しい研究をしてきて「報われた」と感じた経験をしました。
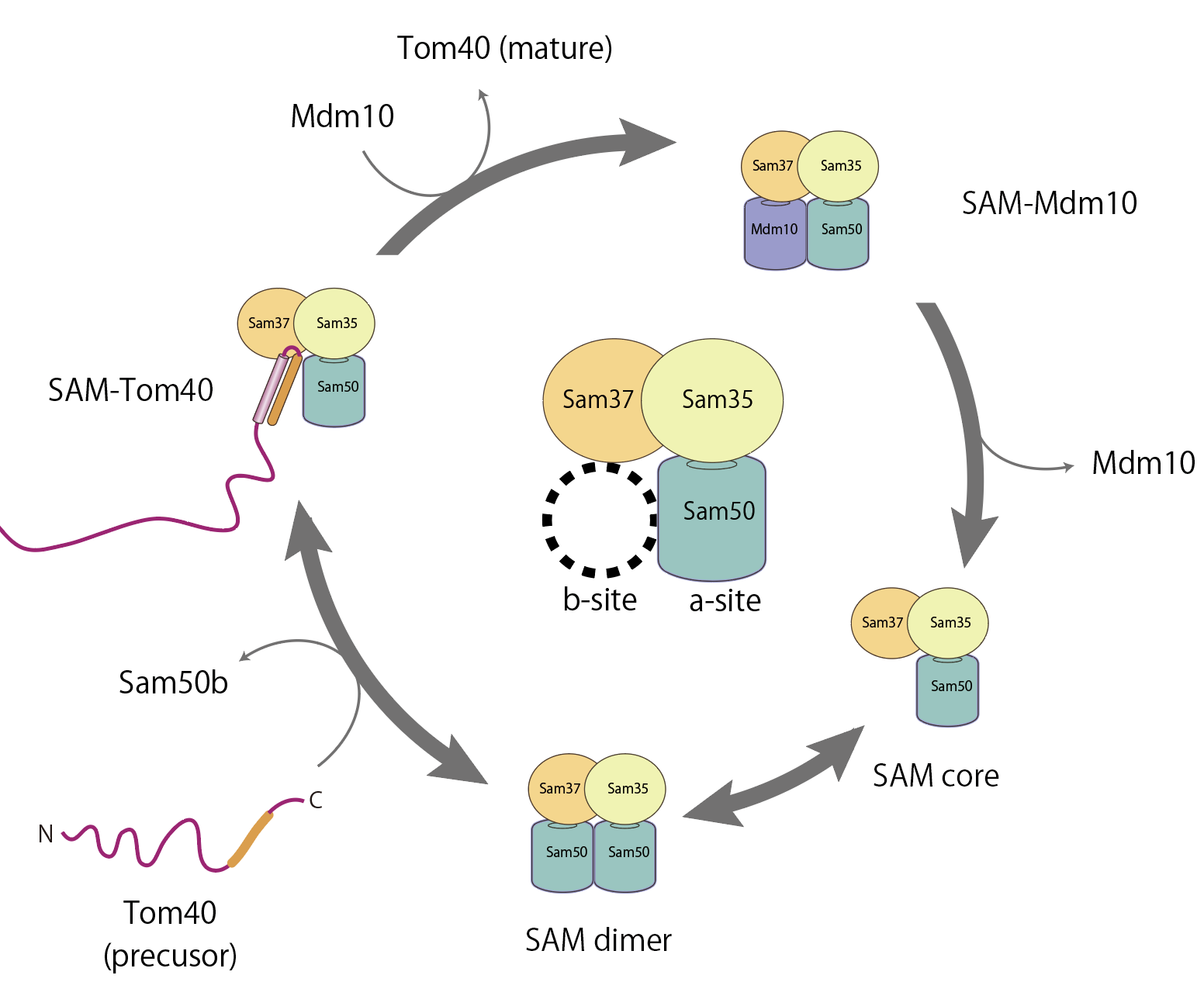
図4:SAM複合体のタンパク質膜挿入におけるb-siteの分子スイッチ機構
しかしこの構造を解いた僅か1か月後に、バクテリア由来のBAM複合体(オルガネラではなく、外膜に存在する膜挿入タンパク質)が基質タンパク質をβバレル型に組み込んでいる構造がNatureで発表されました。種は違えども、自分より先にこの構造を見ている人がいた、しかも論文がNatureにアクセプトされた、と知り、愕然としました。これが構造解析の競争の厳しいところです。私の論文はScienceの査読まで進んだのですが、あと一歩というところで、このBAM複合体の構造が発表され、editorに「もうあなたの解いた構造にインパクトは見出せない」とリジェクトされました。本当に構造解析の分野は競争が激しく、「神様がつくったと思える素晴らしい構造を見ることができる」「しかし競争に負けたら何にもならない」という世界ですね。
聞き手:現在行われているミトコンドリア外膜のご研究について聞かせてください。
現在は、いろんなミトコンドリア外膜のトランスポータタンパク質の構造解析を網羅的に進めています。ミトコンドリア外膜は細胞のエネルギーであるATP合成に最適な環境を作り出すための場です。そのミトコンドリア外膜がどのように成り立っているのか、機能しているのかを統合的に理解したいと思っています(図5)。そのためには、ミトコンドリア外膜にはいろんな膜タンパク質が有りますが、それらを1個1個丁寧に、構造生物学的に機能のメカニズムを解明していきたいと考えています。その先には、ミトコンドリア外膜とは何か?という根源的な理解が得られると期待しています。
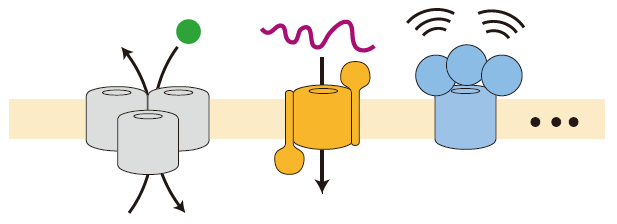
図5:様々なミトコンドリア外膜タンパク質を解明し、ミトコンドリア外膜を統合的に理解
ミトコンドリア外膜の機能を明らかにするということは、分子進化の最終段階を見ることになると思います。ミトコンドリアというのは、元々はバクテリアです。バクテリアも外膜を持っていますが、それが最終的には高等生物の細胞のミトコンドリア外膜まで分子進化しました。今研究しているのは酵母由来のミトコンドリア外膜のタンパク質ですから、その進化の途中を見ているという事になるのかな、と考えています(図6)。私が現在、これからも関わっていきたいのは、そういった進化科学も含めたタンパク質科学です。
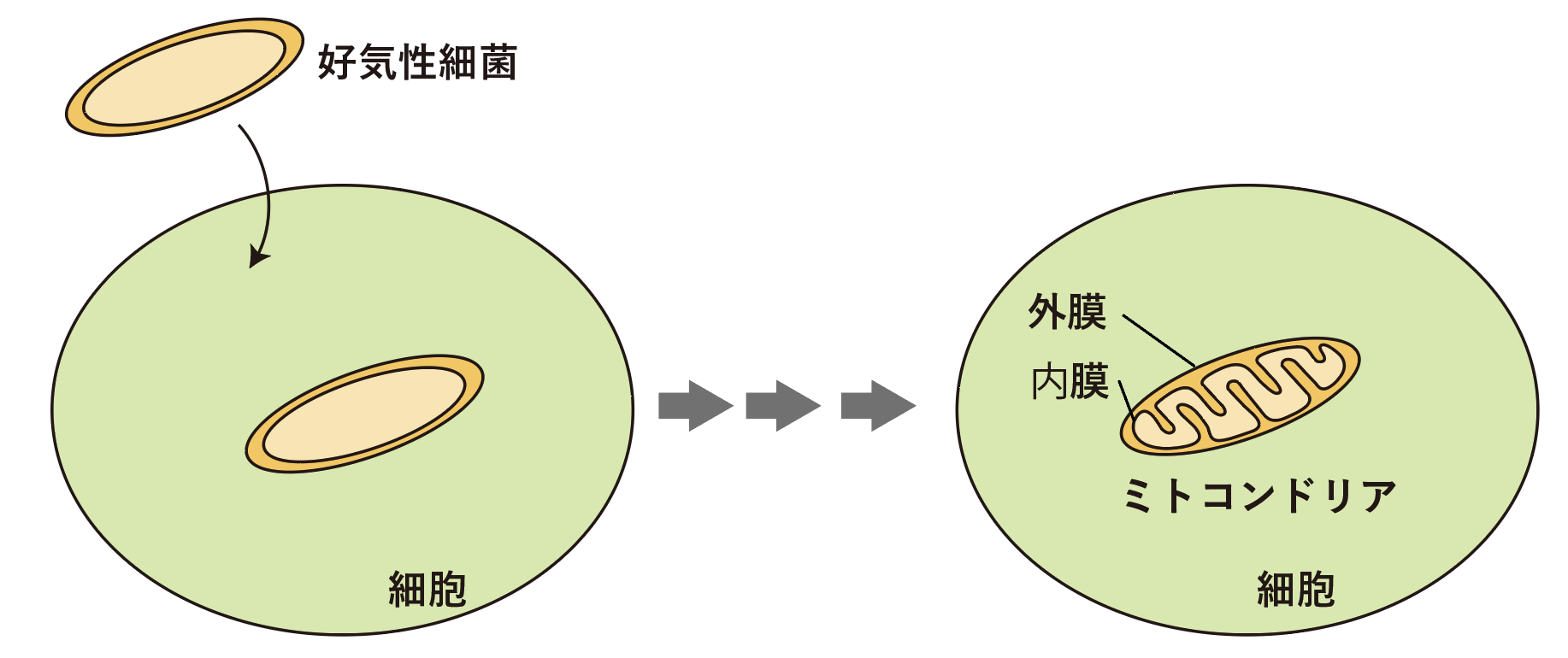
図6:原核生物であるバクテリアの外膜から、真核生物のミトコンドリア外膜としての進化
聞き手:今回、JSTの「創発的研究支援事業」に採択されましたが、今後の意気込みを聞かせてください。
これまで大型予算に沢山申請を出してきましたが、何度も跳ね返されてきました。私自身による研究成果を少しずつ積み上げて、ようやく研究の継続性とか、インパクトが審査員の先生方に伝えられるようになったのかなと思います。今回選んでいただいた厚意に恥じない研究をしていきたいと思います。
これまでの研究の経験で得た、個々のタンパク質に応じて嗅覚を働かせ、戦略を立て、突拍子もないアイデアで局面を切り抜ける、といった能力こそが自分の武器だと自覚しているので、それを継続して研究を進めていきたいと思います。PIになると、自分は手を動かさず、学生やスタッフにどう結果を出してもらうかが大事になると思います。私はこれまで実験ノートとは別に、「どうやって経験し、修正してきたか」の苦労を研究ノートとして、書き留めていて、これが自分の研究の研究室の理念となると思います。そこに書かれた私の持つノウハウをできるだけ単純化、言語化して、学生さんやスタッフにも分かるようにして伝えていくつもりです。実は私は学生時代に英語のディベートサークルに入り、物事を論理的に考える、体系立てる、という頭の使い方を養っていました。その経験は体にしみついていて、今でも役立っていると思うからです。また、ラボのメンバーのモチベーションを常に高め、いかにお金を取ってくるか、「独立した研究者」としての目線でも研究室を引っ張っていきたいと思います。

先端バイオ工学研究センター建物外観 膨大なタンパク質合成、精製作業を行うベンチにて
聞き手:URAの支援についてコメントをお願いします。
創発の面接練習では大変お世話になりました。面接が苦手だという事を自覚し、模擬面接を何度も行ったことで、かなり鍛えられたと思います。URAの皆さんと審査員になって下さった先生のおかげで、与えられた10分間の説明時間内に、発表内容のどこをそぎ落として、どこを重点的に話すか、はっきりさせることができました。この経験は今後のプレゼンでも非常に役に立つと思います。他の先生も面接まで進んだ際は、必ず事前にURAの皆さんと練習をした方が良いと思います。
また、大型研究費に関して、過去に採択された先生の申請書やヒアリング資料をお手本にするのが有効であると思います。言葉の使い方や読み易さ、構成等で、かなり差が出ると思います。本来は場数を踏んで良い申請書が書けるようにならなければいけないのかもしれないのですが、直近で大きな研究費が必要な若手研究者にとっては、やはりお手本が有ると非常に助かります。
関連リンク
● 神戸大学HP
● researchmap

2024年7月(配信) 聞き手、文責:川上勝